
Гайденок Н.Д.,
Багаев Б.М., Чмаркова Г.М.
КрасГАСА, Красноярск
Проблема циклических колебаний численности (биомассы) является одной из интереснейших в динамике популяций. В литературе, посвященной исследованию данного вопроса, предлагается широкий спектр моделей различного математического характера: от интегро - дифференциальных уравнений в частных производных до нелинейных отображений. Ниже будет показано на примере довольно агрегированной модели использование формализма Лагранжа для определения условий, обеспечивающих режимы колебаний.
Обратимся к модели, где рассматривается популяция, имеющую два и более возрастных класса, можно подразделить на два элемента: пополнение и запас, выделяемые на основе статуса половозрелости. Дискретной моделью, описывающей динамику численности популяции, имеющей состав ``пополнение (P) - запас (Z)'' может служить следующая система:
Для удобства выкладок перепишем (2) в виде
Последнее выражение, являющееся уравнением Льенара, можно представить в виде уравнения Лагранжа типа (4):
Временная развертка динамики популяции отражает затухающие колебания вокруг некоторой точки. Однако, в природе у ряда популяций существуют устойчивые колебания численности. Ярким примером таких колебаний могут служить колебания численности тихоокеанских лососевых и, особенно, горбуши. Но, в модели (2.2) построенной на довольно естественных положениях их не воспроизводится на сравнительно длинном временном интервале. В чем здесь причина? Иными словами, возникает довольно формальная задача, заключающаяся в регламентации параметров данной модели (из области допустимых) таким образом, чтобы решение (2.2) имело колебательный режим.
Анализ феноменологии, положенной в основу вывода (2.2) показывает, что в
реальных диапазонах параметров d![]() , V и d
, V и d![]() невозможно
сделать декремент затухания нулевым. Более того, для горбуши величины
параметров V=1 и d
невозможно
сделать декремент затухания нулевым. Более того, для горбуши величины
параметров V=1 и d![]() =1 обеспечивают максимальную скорость
затухания а+с = -2.
=1 обеспечивают максимальную скорость
затухания а+с = -2.
Таким образом, здесь возникает чисто формальный вопрос: ``Каким требованиям должна удовлетворять функция f(Z), чтобы решения (2) имели колебательный характер?''
Единственным решением данного вопроса является функция
генерирующая релаксационные колебания решений расширенной системы, представленные ниже
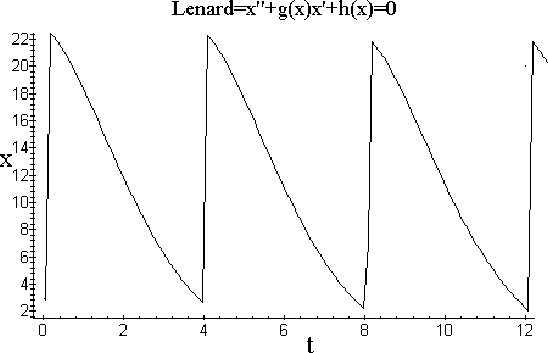
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Saturday, 29-Sep-2001 19:17:34 NOVST