
Суховольский В.Г.
Институт леса им.В.Н.Сукачева СО РАН, Красноярск
It is considered an optimization approach to description of individuals and populations in ecosystems. The approach combines resource and competition paradigms of ecology. There have been introduced definitions of ecological utility function, principle of ecological utility maximum, and ecological prices. Models of food consumption by forest insects and forest insect population dynamics are proposed. The models take into account both population and energy parameters of individuals and host plants.
Ключевые слова: оптимизационный подход, экологическая полезность, модель динамики численности.
В течение последних ста с лишнем лет в экологии был накоплен огромный фактический материал о процессах, происходящих в экосистемах, предложено значительное число теорий, объясняющих те или иные стороны жизнедеятельности и взаимодействия особей и популяций в экосистемах.
Все эти теории основывались на двух основных метатеориях (парадигмах): ресурсной парадигме и парадигме конкуренции.
Согласно ресурсной парадигме процесс роста и развития особей и популяций определяется объемом доступных ресурсов. Недостаток тех или иных ресурсов, требуемых для роста особей, лимитирует процессы функционирования особей. популяций и экосистем. Это положение известно как принцип Либиха.
Другой парадигмой, служащей одной из основ теоретического лесоведения, является парадигма меж- и внутрипопуляционной конкуренции. В биологии эта парадигма идет от Ч.Дарвина, объяснявшего процесс эволюции конкуренцией между популяциями и внутри популяций и выживанием наиболее приспособленных особей, дающих наиболее многочисленное потомство.
Представление о конкуренции особей в популяции и популяций в экосистеме как о причине изменений численностей этих популяций в математической форме было выражено В.Вольтерра и А.Лотка.
Хотя ресурсная и конкурентная парадигмы логически хорошо сочетаются - между особями и популяциями идет борьба за ресурсы и победившие в этой борьбе получают больший объем ресурсов, что способствует росту этих особей и популяций, - в современной экологии существует разрыв между описанием экосистем c точки зрения потоков вещества и энергии и сообществ - с популяционной. Модели динамики численности, начиная с моделей В.Вольтерра, не учитывают энергетику процессов роста, а при описании энергетического баланса в экосистеме не принимают во внимание процессы, связанные с изменением численности популяций в экосистеме. В фундаментальных учебниках по экологии, например в книге Ю.Одума (1986), разделы, посвященные анализу динамики численности популяций и рассмотрению закономерностей циркуляции вещества и энергии в экосистемах, можно изучать совершенно независимо друг от друга.
Возможны ли другие подходы к описанию процессов функционирования популяций в
экосистеме? Достаточно давно различные исследователи предлагают использовать
для описания процессов роста деревьев и древостоев некоторые экстремальные
принципы, подобные принципу наименьшего действия в механике или принципу
кратчайшего пути распространения света в оптике. Наиболее часто в биологии
оптимизационный принцип используется в форме, предложенной Р.Фишером, как
принцип максимума относительной скорости роста ![]() организма или популяции
численностью
организма или популяции
численностью ![]() :
:
Однако биологический смысл подобного экстремального принципа неясен. Представляется, что в данном случае произошло смешение принципа оптимизации функции состояния объекта с принципом оптимизации параметра состояния объекта. Ни один из экстремальных принципов физики не формулируется так, что тот или иной параметр состояния физического объекта стремится к максимуму или минимуму. Второй закон термодинамики утверждает, что в замкнутой системе энтропия, характеризующая функцию распределения физических объектов по определенным параметрам, стремится к максимуму, а вовсе не то, что импульс или скорость этих объектов стремятся к максимуму или минимуму.
И в физике, и в экономике первым свойством оптимизационной метатеории является существование некоторого основополагающего принципа, определяющего поведение и физических объектов, и экономических субъектов. Для физики таким основополагающим принципом является принцип наименьшего действия. Для экономики аналогом принципа наименьшего действия является представление о разумности поведения экономических субъектов. Под разумностью понимается то положение, что экономические субъекты не будут действовать себе во вред или вести себя случайным образом, а будут стремиться вести себя так, чтобы максимизировать свою прибыль или полезность своих затрат.
Вторым свойством оптимизационных теорий является наличие функции, конкретизирующей принцип наименьшего действия или принцип максимизации доходов. Математическая формулировка оптимизационного принципа наименьшего действия и принципа максимизации доходов состоит в том, что физическая система ''стремится'' к состоянию с минимальной энергией, а денежные ресурсы тратятся так, чтобы значение функции дохода или общей функции полезности от купленных товаров было максимально.
Далее, для физической теории необходимо существование функции, описывающей зависимость величины энергии системы от ее параметров, таких как скорость, масса и т.д. Аналогом таких функций в экономике являются функции полезности, характеризующие степень удовлетворенности потреблением того или иного товара в зависимости от предлагаемого объема товара.
Следующим важным принципом, без которого никакая физическая и экономическая теория существовать не могут, является принцип бюджетного сдерживания. В экономике расходы экономических субъектов не могут быть неограниченными. В физике должен выполняться закон сохранения энергии в замкнутой системе.
Еще одним понятием, без которого никакая физическая и экономическая теория не может существовать, является понятие цены. Аналогом цены в физике является энергия взаимодействия. Как невозможно купить товар, если его цена превосходит бюджетные возможности покупателя, так и невозможно физическое взаимодействие, для осуществления которого нужна энергия, превосходящая ту, что имеется у физической системы.
И, наконец, последним принципом, необходимым для построения физической и экономической оптимизационной метатеории, является принцип оптимальности. Принцип оптимальности в той форме, в которой его ввел Р.Беллман, гласит, что оптимальная стратегия обладает следующим свойством: каковы бы ни были начальные условия и первое оптимальное решение, оставшиеся решения должны быть элементами оптимальной стратегии относительно состояния, определенного первым решением.
По аналогии с физикой и экономикой можно говорить о построении экологической метатеории, описывающей функционирование особей, популяций, сообществ и экосистем, которая оперирует с понятиями и принципами, аналогичными соответствующим понятиям и принципам физической и экономической оптимизационных метатеорий.
В такой теории должен существовать принцип, аналогичный принципу наименьшего действия или принципу максимизации доходов.
Далее, должна быть введена функция - аналог функций доходов или полезности в экономике. Рост особи или популяции должен идти так, чтобы эта функция достигала своего максимума или минимума. Естественным образом можно ввести аналог принципа бюджетного сдерживания, так как все процессы в экосистеме ограничены количеством энергии, получаемой в процессе фотосинтеза и объемом других доступных ресурсов (минеральных, водных и др.). Естественно также введение аналогов цен, характеризующих энергетическую стоимость синтеза единицы массы той или иной фракции дерева. Наконец, в какой-либо форме должен выполняться принцип оптимальности, гарантирующий, что любая часть оптимальной траектории, описывающей поведение экосистемы и ее компонентов во времени, также будет обладать оптимальными свойствами.
Разумеется, для различных биологических систем оптимизационные теории и конкретные характеристики оптимизационных моделей могут быть различными. Так, оптимизационный подход широко используется для объяснения стратегий распределения ресурсов в сообществе, размножения и территориального поведения животных. При этом для описания поведения животных (или групп животных) вводится две функции: функция полезности, характеризующая степень удовлетворения объемом добываемых пищевых ресурсов в зависимости от размеров контролируемой особью или группой особей территории и плотности ресурсов на этой территории, и функция расходов энергии на контроль территории в зависимости от ее размеров. При неограниченном росте размеров территории функция полезности стремится к некоторому пределу (связанному с физической невозможностью освоить территорию больше определенных размеров) ограниченном росте размеров контролируемой, а расходы энергии на контроль и защиту территории растут с ростом размеров территории, то должен существовать некоторый оптимальный для данных условий размер территории, обеспечивающий максимальную величину полезной энергии (энергия, получаемая от ресурсов на территории минус энергия, потраченная на освоение и защиту территории), получаемой особью, контролирующей территорию. В данном случае максимизируется не параметр, характеризующий особь размер контролируемой территории, а функция - полезная энергия, получаемая от эксплуатации контролируемой территории. При этом при различной плотности ресурсов и конкурентном давлении размеры территории, контролируемой особью (или группой особей), будут варьировать, но во всех случаях предполагается, что контролируемая территория имеет такие размеры, что обеспечивается выполнение условия максимизации полезной энергии, получаемой в процессе эксплуатации территории.
При анализе распределения ресурсов возможно пользоваться методами, аналогичными применяемым при анализе прибыли и затрат в экономике, причем прибыль в экологии это повышение приспособленности и увеличение вероятности выживания, а затраты энергия и время, необходимые для обеспечения размножения.
По аналогии с экономическим подходом рассмотрим модель потребления корма
гусеницами насекомых - филлофагов. Все метаболические процессы разделим на
две основных группы - связанные с усвоением корма личинкой насекомого
![]() , и метаболические процессы, связанные с синтезом ее биомассы
, и метаболические процессы, связанные с синтезом ее биомассы
![]() и поддержанием ее существования. Тогда балансовое уравнение
энергетики потребления корма насекомыми имеет вид:
и поддержанием ее существования. Тогда балансовое уравнение
энергетики потребления корма насекомыми имеет вид:
При построении модели потребления корма гусеницами допустим, что
метаболическая энергия, потраченная на обеспечение роста и существования
особи, прямо пропорциональна энергии, аккумулированной в биомассе гусеницы:
![]() , а метаболическая энергия, потраченная на усвоение корма
личинкой насекомого, задается соотношением
, а метаболическая энергия, потраченная на усвоение корма
личинкой насекомого, задается соотношением
![]() . Тогда из
уравнения (1) после простых преобразований можно получить уравнение
. Тогда из
уравнения (1) после простых преобразований можно получить уравнение
В плоскости
![]() уравнение (2) есть прямая, пересекающая ось
абсцисс в точке
уравнение (2) есть прямая, пересекающая ось
абсцисс в точке ![]() , а ось ординат - в точке
, а ось ординат - в точке ![]() . Уравнение (2) можно трактовать
как прямую бюджетных ограничений, а величины (
. Уравнение (2) можно трактовать
как прямую бюджетных ограничений, а величины (![]() и
и ![]() - как ''цены'' синтеза
биомассы и усвоения корма. Предположим по аналогии, что максимальная
полезность потребления корма, которую можно трактовать как минимальный риск
гибели особи, достигается в точке на плоскости
- как ''цены'' синтеза
биомассы и усвоения корма. Предположим по аналогии, что максимальная
полезность потребления корма, которую можно трактовать как минимальный риск
гибели особи, достигается в точке на плоскости ![]() , в которой
произведение
, в которой
произведение ![]() достигает максимума:
достигает максимума:
Решение оптимизационного уравнения (3) дает оптимальную для данных ''цен''
корма величину коэффициента использования корма на синтез биомассы особи
![]()
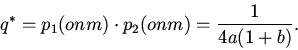
Значения коэффициентов уравнений связи параметров ![]() и
и ![]() ,
полученные по данным экспериментов по выкармливавшимся на хвое
лиственницы сибирской самцам и самкам непарного шелкопряда, хорошо
согласуются с уравнениями (1) - (3).
,
полученные по данным экспериментов по выкармливавшимся на хвое
лиственницы сибирской самцам и самкам непарного шелкопряда, хорошо
согласуются с уравнениями (1) - (3).
Таким образом, энергетическую эффективность питания гусениц определяют две
''цены'' - ''цена'' усвоения корма и ''цена'' синтеза биомассы особи. Из
этого следует, что при фиксированном значении ![]() эффективности
питания возможны две предельные стратегии питания. Первая стратегия состоит
в использовании корма с высокой ''ценой'' усвоения, но низкой ``ценой''
синтеза биомассы гусениц. Вторая - противоположная - стратегия состоит в том,
чтобы использовать корм с низкой ''ценой'' его усвоения насекомым, но
высокой ''ценой'' синтеза биомассы особей. Увеличение же экологической
эффективности питания гусениц будет связано с выбором корма с низкими
''ценами'' усвоения и синтеза биомассы.
эффективности
питания возможны две предельные стратегии питания. Первая стратегия состоит
в использовании корма с высокой ''ценой'' усвоения, но низкой ``ценой''
синтеза биомассы гусениц. Вторая - противоположная - стратегия состоит в том,
чтобы использовать корм с низкой ''ценой'' его усвоения насекомым, но
высокой ''ценой'' синтеза биомассы особей. Увеличение же экологической
эффективности питания гусениц будет связано с выбором корма с низкими
''ценами'' усвоения и синтеза биомассы.
Использование понятий энергетической ''цены'' дает возможность говорить о
реакции особей насекомых на изменения характеристик кормовых растений. Так,
уменьшение антибиозных свойств листьев приводит к уменьшению ''цены''
усвоения корма. Тогда при сохранении неизменной ''цены'' синтеза биомассы
гусениц точка пересечения прямой бюджетных ограничений с осью ![]() сдвинется ближе к нулю. Это приведет к соответствующему уменьшению величины
сдвинется ближе к нулю. Это приведет к соответствующему уменьшению величины
![]() , то есть к уменьшению относительной биомассы особей, уменьшению
плодовитости самок и другим внутрипопуляционным перестройкам. Развитие этого
подхода позволило построить оптимизационную модель динамики численности
популяций лесных насекомых, в которой учитываются как популяционные, так и
энергетические аспекты функционирования популяций в экосистеме.
, то есть к уменьшению относительной биомассы особей, уменьшению
плодовитости самок и другим внутрипопуляционным перестройкам. Развитие этого
подхода позволило построить оптимизационную модель динамики численности
популяций лесных насекомых, в которой учитываются как популяционные, так и
энергетические аспекты функционирования популяций в экосистеме.
Из предложенной модели следует, что возможны вспышки массового размножения двух типов. Вспышки первого типа связаны с уменьшением массы особей в популяции в процессе развития вспышки. Вспышки второго типа характеризуются увеличением массы особей в ходе вспышки. Эти положения модели подтверждаются наблюдениями за популяциями лесных насекомых, дающих вспышки массового размножения.
Для ряда видов лесных насекомых (например, для сосновой пяденицы Bupalus piniarius L. и для непарного шелкопряда Limantria dispar L. развитие градации популяции сопровождается уменьшением массы куколок, причем это уменьшение наблюдается тогда, когда еще нет каких-либо сильных повреждений хвои и нельзя говорить о недостатке корма.
У сибирского шелкопряда Dendrolimus superans sibiricus Tschetv в процессе развития вспышки массового размножения этого вида масса отдельных особей увеличивается.
В этом случае вспышка массового размножения сибирского шелкопряда должна быть связана с уменьшением цены корма, а вспышка массового размножения непарного шелкопряда - с увеличением цены корма.
Одновременное рассмотрение как популяционных, так и энергетических характеристик особей в популяции позволяет описать возникновение вспышек массового размножения, учесть эффекты влияния как модифицирующих, так и регулирующих факторов на изменение численности популяции, учесть эффекты антибиоза и потери устойчивости кормовых растений.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Tuesday, 04-Sep-2001 18:33:08 NOVST