Фазовые портреты и методы главных и гладких компонент в анализе динамики численности и структуры популяций
Ефимов В.М.,
Ковалева В.Ю.
Институт систематики и экологии животных СО РАН
Аннотация:
Совокупность отрезков длины одномерного временного ряда можно
рассматривать как точки -мерного фазового пространства. Соединяя их
последовательно, например сплайнами, получим траекторию ряда в этом
пространстве - многомерный фазовый портрет. Для редукции размерности можно
применить метод главных компонент, заключающийся в поиске координатных осей,
в проекции на которые дисперсия траектории ряда максимальна. Максимизация
автоковариации вместо дисперсии приводит к методу гладких компонент. Оба
метода применимы к любому временному ряду, не требуют его стационарности и
оказываются исключительно полезны при анализе внутренних закономерностей и
прогнозе динамики численности и структуры популяций животных и влияющих на
них факторов.
Изложены результаты применения этих методов к анализу динамики
численности и структуры популяции полевки-экономки в Горном Алтае.
Set of one-dimensional time series segments of length may be consider as a
set of points in -dimensional phase space. Series trajectory in this space
- multidimensional phase portrait - is a consecutive join of this points,
for example, by splines. The principal components method is suitable for
reduction of dimension. It consist in founding of coordinate axes, on which
the series trajectory projection variance is maximal. Maximization of
autocovariance instead of variance lead to the smooth components method.
Both methods are applicable to any time series without stationary
requirement and very useful for analysis and prediction of animal population
structure and density dynamics and influencing factors. Results of this
methods application to root vole population density and structure dynamics
in Mountain Altai are presented.
Ключевые слова: Фазовый портрет, методы главных и гладких компонент,
прогноз динамики численности.
Первому автору посчастливилось слушать лекции Алексея Андреевича Ляпунова в
физико-математической школе при НГУ и самом НГУ в 1964-70 годах. В конце
этого периода, конечно же при содействии ''Факела'', была проведена научная
конференция студентов и школьников и после нее Алексей Андреевич пригласил
всех участников, в докладах которых были хоть какие-нибудь попытки применить
математику в биологии, на беседу за чашкой чая к себе в коттедж, уделив
каждому, несмотря на всю свою занятость, отдельный вечер. Я думаю, что
сейчас такое даже трудно себе представить. Поразило его вдумчивое и
серьезное отношение к достаточно частной и, казалось бы, далекой от его
интересов работе. Темой беседы являлись методы учетов численности животных и
так уж случилось, что основная часть моей последующей профессиональной
деятельности оказалась связанной с динамикой популяций и компьютерной
обработкой данных по учетам численности животных.
Пусть имеется последовательность
наблюдений некоторого
показателя в равноотстоящие моменты времени . Пусть
где - число лет наблюдений, - лаг. Пусть уже центрирована и нормирована, и - матрица собственных векторов . Умножая на
, получим - матрицу главных
компонент [4].
Обозначим через матрицу без первой строки, через - матрицу
без последней строки. Тогда
Так как матрица симметрична, для его решения достаточно
применить стандартный метод нахождения собственных векторов и значений. В
результате получим
где - ортогональная матрица собственных векторов,
- диагональная матрица собственных значений.
Умножая на , получим - матрицу гладких компонент [3]. В отличие от главных компонент, которые всегда ортогональны друг другу, гладкие компоненты не обязаны
быть ортогональными, несмотря на ортогональность , и являются,
таким образом, разновидностью факторного анализа. Однако на практике
корреляции между ними обычно невелики, что позволяет относиться к ним как
достаточно независимым составляющим матрицы . Второе
отличие заключается в том, что собственные значения могут быть
отрицательными, если отрицательны соответствующие автоковариации.
Откладывая на плоскости точки
,
где -тый столбец матрицы , и
соединяя их последовательно, получим фазовые траектории временного ряда на
плоскости -той и -той главных или гладких компонент. Эти методы
применимы к любому временному ряду, не требуют
его стационарности, автоматически выявляют тренд и оказываются исключительно
полезны при анализе внутренних закономерностей и прогнозе динамики
численности и структуры популяций животных и влияющих на них факторов.
Проблеме популяционных циклов животных посвящена огромная литература, однако
она до сих пор далека от разрешения.
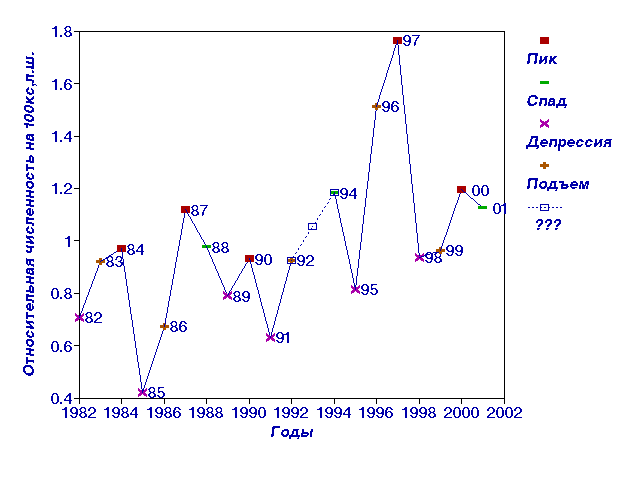
Рис. 1.
Динамика относительной численности полевки-экономки Прителецкой
тайги.
На рис. 1 изображена динамика относительной численности полевки-экономки
Прителецкой тайги (Microtus oeconomus) c 1982 по 2001 год, для которой, так же как и для
других лесных полевок, характерны популяционные циклы с периодом 3-4 года и
отмечены фазы этих циклов. Данные за 1993 год отсутствуют. Более или менее
очевидно, что 1992 год является годом подъема. Что касается 1994 года, то он
может быть как годом пика, так и годом спада.
Чтобы решить этот вопрос, в 1993 год было подставлено среднее между 1992 и
1994 годом и полученный ряд был интерполирован сплайнами и обработан методом
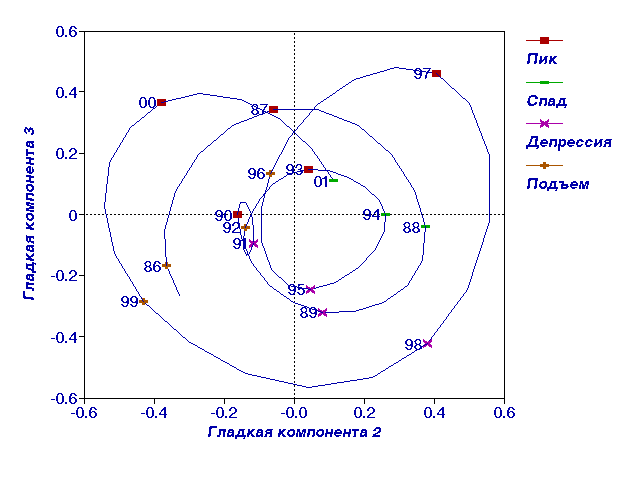
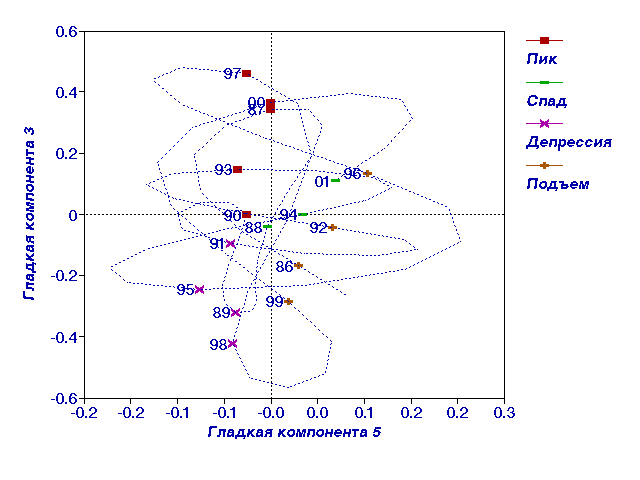
гладких компонент с лагом 4 года (рис.2, 3). В первой компоненте проявился
тренд с примерно 9-10 летними колебаниями, а во второй и третьей -
3-4-летние колебания (рис. 2). Из рисунков видно, что наиболее естественно
считать 1993 год годом пика, а 1994 год - годом спада численности. На рис.
3 все годы, относящиеся к каждой из фаз, расположились рядом друг с другом и
маркируют часть плоскости, относящейся к каждой фазе, позволяя довольно
уверенно отнести нерасклассифицированные годы к той или иной фазе.
Рис. 2.
II и III гладкие компоненты динамики численности полевки-экономки.
Рис. 3.
V и III гладкие компоненты динамики численности полевки-экономки.
Несмотря на многочисленные исследования, до сих пор не имеет однозначного
решения и вопрос о взаимосвязи морфологической и физиологической
изменчивости с плотностью популяции.. И гипотеза Кристиана [2] и гипотеза
Читти [1] предсказывают, хотя и на разной основе, циклические изменения
фенооблика популяции в соответствии с динамикой численности. Проверка такого
предсказания на природных популяциях требует долговременных рядов
наблюдений. Однако, к сожалению, в большинстве работ длительность изучаемых
рядов не превышает одного-двух популяционных циклов.
Материалом для исследования послужили выборки из популяции полевки-экономки
(Microtus oeconomus Pallas 1776) черневой низкогорной тайги (Республика Алтай, Турочакский
район, с.Артыбаш), взятые в 1989-92, 94-2000 годах. За это время в популяции
прошло почти четыре цикла динамики численности (рис.1). Были вычислены
отдельно по самцам и самкам годовые средние длины тела, хвоста и ступни,
веса тела, селезенки, правого надпочечника и правого семенника (для самцов)
и исследована динамика как самих показателей, так и разности по ним между
самцами и самками.
Наиболее четко циклические изменения, соответствующие динамике численности
популяции, проявились у самок по весу тела и надпочечника, а у самцов - по
весу надпочечника, семенника и селезенки. Однако у самок максимальные
значения этих показателей приходятся на годы пиков, а у самцов, в отличие от
самок, запаздывают на год и попадают на фазу спада или депрессии. Поэтому
динамики годовых средних были обработаны методом главных компонент отдельно
для самцов и самок. Этим же методом обрабатывались динамики разностей
годовых средних между самками и самцами (рис.4). Во всех трех случаях на
первую компоненту пришлось от 63 до 69% , на вторую - от 14 до 22% .
Структура собственных векторов также оказалась одинаковой. В первую
компоненту внесли положительные вклады все признаки, за исключением длины
ступни, которая образовала вторую компоненту. Интерпретация первой
компоненты очевидна - общие размеры особей.
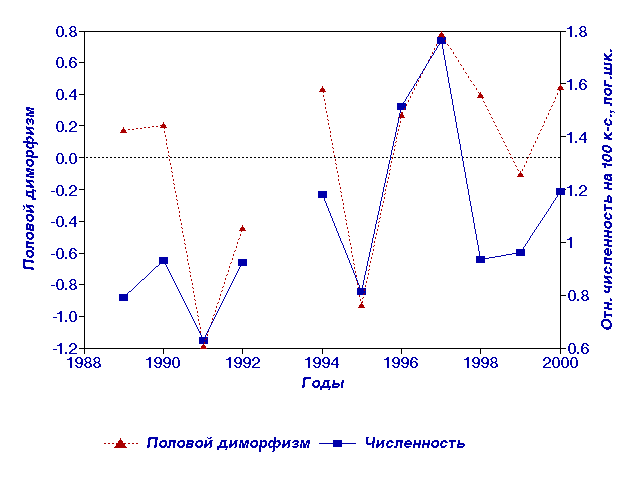
Рис. 4.
Плотностно-зависимая динамика первой компоненты полового диморфизма
полевки-экономки по морфофизиологическим признакам.
Корреляция размерных компонент с логарифмом относительной численности
оказалась незначимой и для самок и для самцов . Однако
первая компонента полового диморфизма - разницы в общих размерах -
достоверно
коррелирует с динамикой численности, практически
полностью повторяя ее очертания (рис.4). Чем выше плотность популяции, тем
сильнее относительное угнетение развития самцов по сравнению с самками.
Таким образом, многомерный анализ позволил выявить плотностно-зависимую
динамику полового диморфизма полевки-экономки по совокупности
морфофизиологических признаков. Аналогичный по содержанию результат был
получен ранее Ю.К.Галактионовым на водяной полевке.

![\begin{displaymath}
r=\sum_{t=1}^{N-1}[\sum_{j=1}^{M}(a_{j}x_{tj})\sum_{j=1}^{M}a_{j}x_{(t+1)j})]
=(X_{N}a)'X_{1}a=a'X_{N}'X_{1}a=a'Va,
\end{displaymath}](img14.gif)

![[SBRAS]](/gif/s_sigma.gif)