Непрерывно-дискретная модель динамики системы "хищник-жертва".
Волкова Е.В. Новосибирский государственный университет
Аннотация:
Работа посвящена построению непрерывно-дискретной модели динамики системы
"хищник-жертва". В работе проводится анализ математической модели динамики
численности взаимодействующих популяций, в рамках которой предполагается,
что появление особей новых генераций происходит в некоторые фиксированные
моменты времени ,
, а смертность
носит непрерывный характер. С помощью численного анализа изучены
динамические режимы изменения численности популяций и показано, что могут
возникать циклические режимы, с циклами различных длин.
Abstract:
This work is devoted to the construction of the discrete-continuous model of
the "predator - prey" system dynamics. We suppose, that the new
generation individuals appearance occurs in the some fixed time point ,
, and the mortality is continuous. Using
the numerical analysis, the dynamic conditions of the interactive
populations population changing are examined and showed, that the cycles of
the different lengths can arise in the model.
Описание модели.
Одним из основных элементов в построении математической модели динамики
численности популяции является учет баланса в изменении соотношений
численностей различных групп в структуре популяции под воздействием
каких-либо факторов. В нашем случае мы предполагаем, что баланс численности
популяции складывается из следующих основных процессов: рождаемости,
процесса саморегуляции, естественной смертности, а также гибели жертв при
столкновении с хищником.
Дополнительно будем предполагать, что численность популяции является
непрерывной величиной. Мы рассмотрим математическую модель динамики
численности изолированной популяции, в которой смертность особей носит
непрерывный характер и каждая особь может погибнуть (под воздействием
различных факторов) в любой момент времени, а увеличение численности
(появление особей новых генераций) происходит в некоторые фиксированные,
равноотстоящие друг от друга моменты времени
На каждом временном интервале
динамику численности
жертв в наиболее простом случае можно описать уравнением, аналогичным
уравнению Ферхюльста:
(1)
где - коэффициент естественной смертности жертв; - коэфицент саморегуляции. Целесообразно ввести в модель дополнительную
переменную , соответствующую суммарному количеству пищи, находящейся в
пищеварительной системе (ПС) хищников. Пусть - объем ПС одного
хищника. Тогда в каждый момент времени должно выполняться неравенство
(2)
Уровень заполненности ПС хищника можно определять как отношение величины
к текущему максимальному значению (что, однако, не совсем
удобно при численном исследовании поведения решений системы вблизи начала
координат) или же как разность
Естественно полагать, что чем
выше заполненность ПС хищника, тем ниже интенсивность потребления жертв.
Таким образом величина
определяется степенью сытости хищников.
Динамику численности хищников на интервале
можно
описать с помощью уравнения
(3)
где коэффициент описывает процессы саморегуляции численности
популяций хищников, прямо пропорционален числу столкновений
особей между собой, что приводит, например, к распространению болезней,
конкурентной борьбе и т.д., а коэффициент описывает процесс
естественной смертности хищников в предположении, что
меняется линейно по отношению к
(интенсивность гибели голодных хищников несомненно выше, чем у сытых):
(4)
Здесь
максимальная интенсивность гибели (достигается
при
),
минимальная интенсивность гибели (достигается при
.
Динамика наполненности ПС хищника определяется тремя процессами: потоком
корма (за счет потребления жетрв), естественным оттоком, а также оттоком,
связанным с гибелью хищников. Увеличение значения переменнойпроисходит со
скоростью
, где
,
,
,
скорость естественного оттока корма из системы в первом приближении можно
считать пропорциональной величинес некоторым постоянным коэффициентом
За время
в популяции хищников гибнет
особей; с каждой погибшей особью из системы
удаляется
пищи; следовательно, суммарные потери
пищи составляют
. Таким образом,
справедливо следующее уравнение:
(5)
Объединяя соотношения (1) - (5), получаем модель динамики системы
хищник-жертва на интервале
. Обозначим через и численности особей жертв и хищников, доживших до
момента размножения , через и - величины
плодовитости особей (среднее число потомков, приходящихся на одну выжившую
особь). Таким образом, численности новых генераций жертв и хищников
определяются выражениями:
(6)
Достаточно сложным и неясным остается вопрос об изменении величины при
появлении особей новой генерации. Очевидно, что потери биомассы при этом
неизбежны и в наиболее простом случае можно считать, что
где параметр
удовлетворяет соотношению
Анализ модели.
Изучение модели дало нам следующие свойства.
При неотрицательных начальных данных решение системы (1)-(5)
неотрицательно.
Траектории системы (1)-(5) не выходят за границы компакта
хx, где
Получены достаточные условия глобальной устойчивости нуля (достаточные условия вырождения популяции).
Достаточные условия неустойчивости нуля.
Замечание. Относительно можно отметить, что при
отсутствии жертв, т.е. полагая в исходной системе , можно подобрать
коэффициенты так, что будут выполнены достаточные условия неустойчивости
нуля и функция не будет стремиться к нулю. С учетом смысловой нагрузки
переменных это означает, что данная популяция жертв не является единственной
кормовой базой хищников и что при отсутствии жертв хищники могут перейти на
другую, второстепенную кормовую базу и не вымереть. Т.о. вполне обосновано
ввести дополнительное условие на функцию
:
при любых
(заметим, что это условие никак не изменяет результаты, полученные ранее).
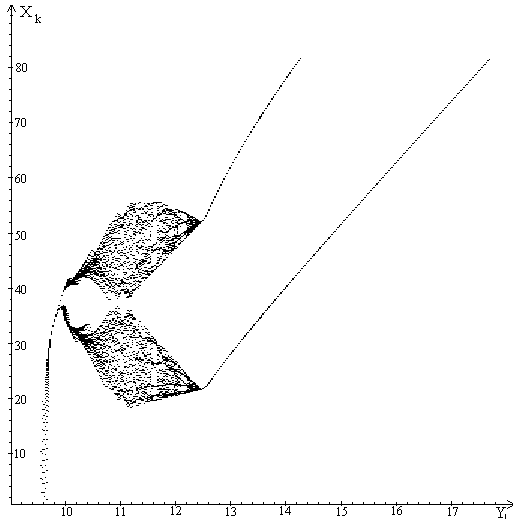
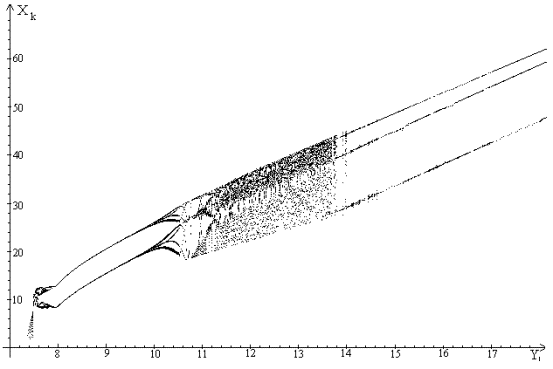
Бифуркационная диаграмма.
Численный анализ модели показал, что модель обладает достаточно богатым
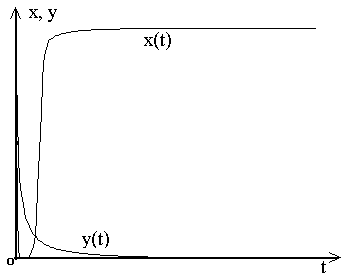
набором динамических режимов. Кроме "обычных " режимов (вырождения обеих
популяций, вырождения хищников при монотонной стабилизации жертв на
ненулевом уровне(рис.1)) наблюдались, в частности, и различные циклические
режимы изменения численностей популяций (рис. 2-5).
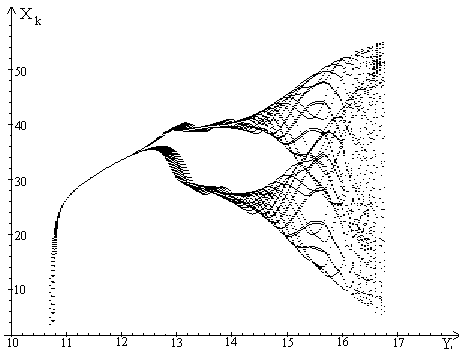
Для наглядного представления динамических режимов, которые могут
реализоваться в рамках нашей модели мы используем бифуркационную диаграмму.
По оси ординат будем откладывать значения координат устойчивых аттрактеров
, а по оси абсцисс - значения параметра , остальные значения
параметров фиксированы.
Рис. 1.
Вырождения хищников при монотонной стабилизации жертв на ненулевом уровне.
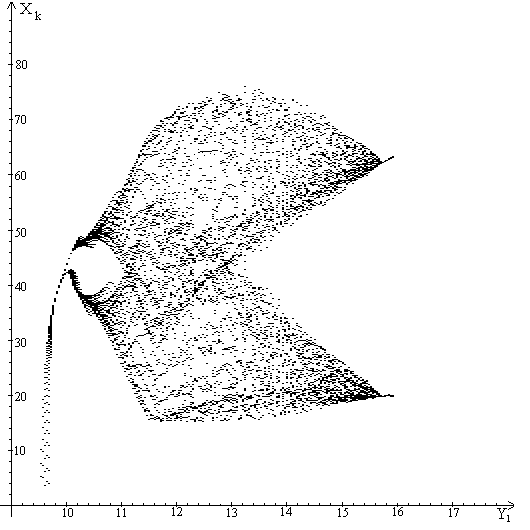
Анализ бифуркационной диаграммы показывает, что у модели (1)-(5) существуют
циклы разной длины (рис. 2-5). Увеличение параметра
приводит к тому, что стационарная точка (0,0,0) теряет устойчивость
и появляется устойчивое ненулевое равновесие. При дальнейшем увеличении
параметра эта точка теряет устойчивость и приводит к
появлению циклов различной длины.
На рис.5 четко видно появление циклов длины три. Известно следствие теоремы
Шарковского о том, что если у отображения действительной полуоси в себя
существует цикл длины 3, то у этого отображения существуют циклы всех длин.
Выводы.
Таким образом, в модели выявлены следующие режимы динамики численности:
вырождение популяции;
стабилизация численности популяции на ненулевом уровне;

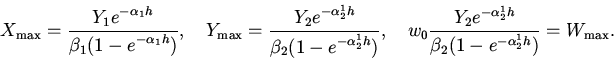

![[SBRAS]](/gif/s_sigma.gif)