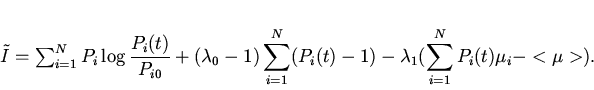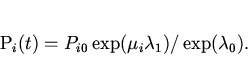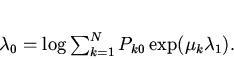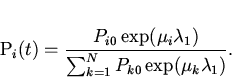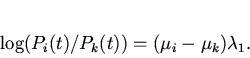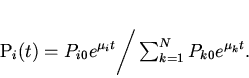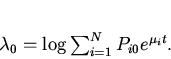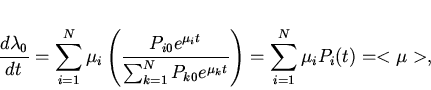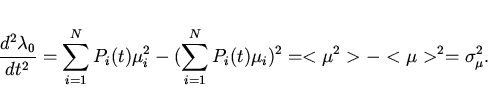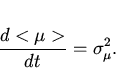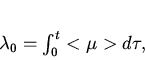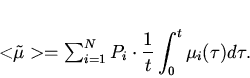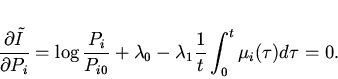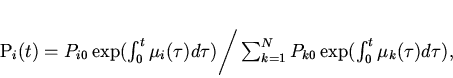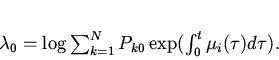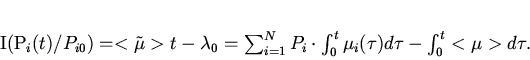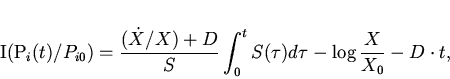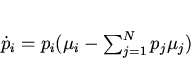 |
(1) |

Евдокимов Е.В.,
Шаповалов А.В.
Томский государственный университет
A variation principle is proposed to describe micro-evolution dynamics of flow haploid populations in the framework of the Janes statistical formalism. The selection dynamics and information production of the population genetic structure are presented in explicit form for the systems under consideration. Biological meaning of basic elements of the variation principle is considered.
Изучение микроэволюционных процессов в биологических популяциях
предполагает описание изменений в их генетической структуре в
ходе отбора. По степени перераспределения генных частот судят об
интенсивности этих процессов. Однако в большинстве случаев
прямое измерение генных частот невозможно по объективным причинам
в силу технических сложностей получения информации на уровне
нуклеотидных последовательностей в ДНК. При экспериментальных
исследованиях как на молекулярном, так и на клеточном уровнях
возникает проблема неполноты их описания с помощью моделей Фишера
- Эйгена [1]
Традиционный подход к решению данной проблемы состоит в использовании измеряемых макропараметров популяции, отражающих ход эволюционного процесса, например среднего значения признака, по которому идет отбор. М. Эйгеном было предложено оценивать скорость микроэволюционных процессов по производству информации, связанной с перераспределением генных частот, посредством информационной меры Кульбака [4,6]. В этой связи представляет интерес теоретический анализ микроэволюционных процессов с использованием меры Кульбака и практических методов ее измерения на основе макропараметров. Одним из известных подходов такого рода является формализм Джейнса, основанный на эвристическом "принципе максимального правдоподобия" предсказания поведения системы при недостатке информации о ней. Согласно Джейнсу [6] наименее предубежденный (наименее ошибочный) способ нахождения распределения вероятностей системы тот, который максимизирует (целевую) функцию, выражающую неопределенность системы при условиях имеющейся информации ( "принцип максимальной неопределенности"). В этом подходе формулируется вариационный принцип максимизации шенноновской меры неопределенности, информационной энтропии рассматриваемой системы. Варьирование информационной энтропии, при наличии уравнений связи между вероятностями различных состояний системы и ее измеряемыми макропараметрами типа средней энергии, дает в итоге функцию распределения вероятностей в явном виде [7,8].
Адаптируем "принцип максимального правдоподобия" для описания эволюции популяций. Поскольку речь идет не о равновесном состоянии системы, а о процессе отбора, модифицируем формализм Джейнса следующим образом.
В соответствии с общим подходом потребуем, чтобы процесс эволюции популяций протекал наименее неожиданным способом.
Рассмотрим вначале эволюцию более простых систем -- турбидостатных
гаплоидных популяций, соответствующих дарвиновским системам с
постоянной организацией и постоянными мальтузианскими параметрами.
Множество генотипов особей, составляющих популяцию, обозначим
![]() , а множество соответствующих им фенотипов (мальтузианских
параметров) через
, а множество соответствующих им фенотипов (мальтузианских
параметров) через ![]() . Положим, что отдельные дискретные
генотипы
. Положим, что отдельные дискретные
генотипы ![]()
![]() однозначно
определяют значения фенотипов, мальтузианских параметров
однозначно
определяют значения фенотипов, мальтузианских параметров
![]() , для носителей данных
генотипов (здесь
, для носителей данных
генотипов (здесь ![]() -- как и ранее, число генотипов в
системе). Это требование естественно для популяций с неполовым
размножением. При случайном выборе какой-либо особи из
исследуемой популяции в момент времени
-- как и ранее, число генотипов в
системе). Это требование естественно для популяций с неполовым
размножением. При случайном выборе какой-либо особи из
исследуемой популяции в момент времени ![]() ее фенотип,
мальтузианский параметр, заранее не известен. Определим его как
случайную величину
ее фенотип,
мальтузианский параметр, заранее не известен. Определим его как
случайную величину ![]() , принимающую дискретные значения
, принимающую дискретные значения ![]() из множества
из множества ![]() . Вероятность того, что случайно выбранная в
момент
. Вероятность того, что случайно выбранная в
момент ![]() особь имеет мальтузианский параметр
особь имеет мальтузианский параметр ![]() ,
определим как
,
определим как
Поставим задачу -- найти вид функции
![]() , используя информацию о
, используя информацию о ![]() и
и ![]() , исходя из
вариационного подхода, подобного формализму Джейнса.
, исходя из
вариационного подхода, подобного формализму Джейнса.
Определим вид варьируемой функции. В ходе эволюции турбидостатной
популяции в результате отбора происходит изменение частот генотипов и
соответствующей им функции распределения,
![]() . Поэтому вместо применяемого Джейнсом для описания
равновесных состояний информационной меры Шеннона в данном подходе
необходимо использовать другую информационную меру, отражающую
ожидаемую неопределенность результатов отбора, или, по терминологии
Реньи [9], неожиданность протекания процесса эволюции. В качестве
меры неожиданности
. Поэтому вместо применяемого Джейнсом для описания
равновесных состояний информационной меры Шеннона в данном подходе
необходимо использовать другую информационную меру, отражающую
ожидаемую неопределенность результатов отбора, или, по терминологии
Реньи [9], неожиданность протекания процесса эволюции. В качестве
меры неожиданности ![]() -го события в теории информации [10]
используется величина
-го события в теории информации [10]
используется величина ![]() , где
, где ![]() - вероятность этого
события. Соответственно, изменение неожиданности для
- вероятность этого
события. Соответственно, изменение неожиданности для ![]() -го генотипа в
ходе отбора можно определить как
-го генотипа в
ходе отбора можно определить как
![]() .
Усредняя по всей популяции в момент времени
.
Усредняя по всей популяции в момент времени ![]() , получим функцию
неожиданности эволюции популяции в виде:
, получим функцию
неожиданности эволюции популяции в виде:
Таким образом, декларируемый в данной работе принцип минимальной неожиданности протекания эволюции идентичен принципу минимального производства информации в ходе отбора или минимальной скорости прохождения системы по координате информационного расстояния. Здесь под информацией понимается информация о функции распределения генотипов в популяции.
Сформулируем
соответствующий вариационный принцип следующим образом:
Функцию ![]() в соответствии с
подходом Джейнса найдем методом неопределенных множителей Лагранжа.
Обозначим множители Лагранжа через
в соответствии с
подходом Джейнса найдем методом неопределенных множителей Лагранжа.
Обозначим множители Лагранжа через ![]() и
и ![]() и введем
функцию Лагранжа
и введем
функцию Лагранжа
Рассмотрим биологический смысл ![]() . Величина
. Величина ![]() безразмерна. Дифференцируя (12) по времени, получим
безразмерна. Дифференцируя (12) по времени, получим
Следствием выражений (13) и (14) является известная
"основная теорема естественного отбора" Фишера , связывающая дисперсию
мальтузианского параметра в популяции и интенсивность отбора,
измеряемую через скорость изменения его среднепопуляционного значения:
Полезным для экспериментальной практики является следствие из
(13) вида
На основе полученных
соотношений можно решить поставленную вначале работы задачу --
количественно оценить производство информации в ходе эволюции
турбидостатной популяции. Из выражений (4) и (11) получаем
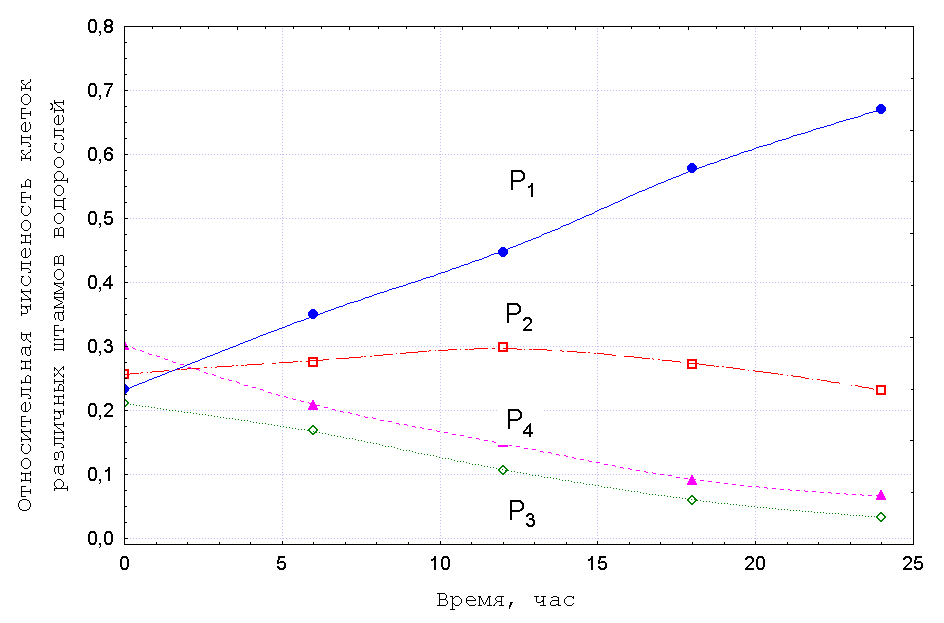
Проиллюстрируем этот подход экспериментом, выполненным Л. Н. Цоглиным с
сотрудниками, в котором изучали ход отбора разных штаммов
микроводорослей при проточном культивировании в турбидостате [12].
На Рис.1 представлены изменения относительных концентраций
клеток разных штаммов, ![]() , прямо измеренные в ходе опыта с помощью
камеры Горяева под микроскопом.
, прямо измеренные в ходе опыта с помощью
камеры Горяева под микроскопом.
 |
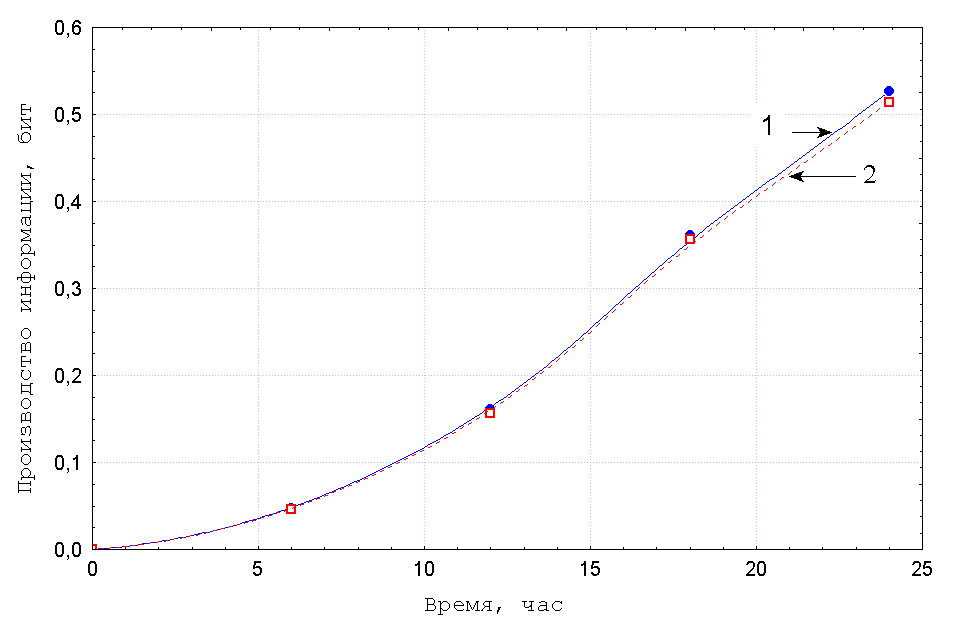 |
Как видно из рисунка, указанные кривые совпадают с точностью до ошибки измерений, что свидетельствует об адекватности предлагаемого нами подхода.
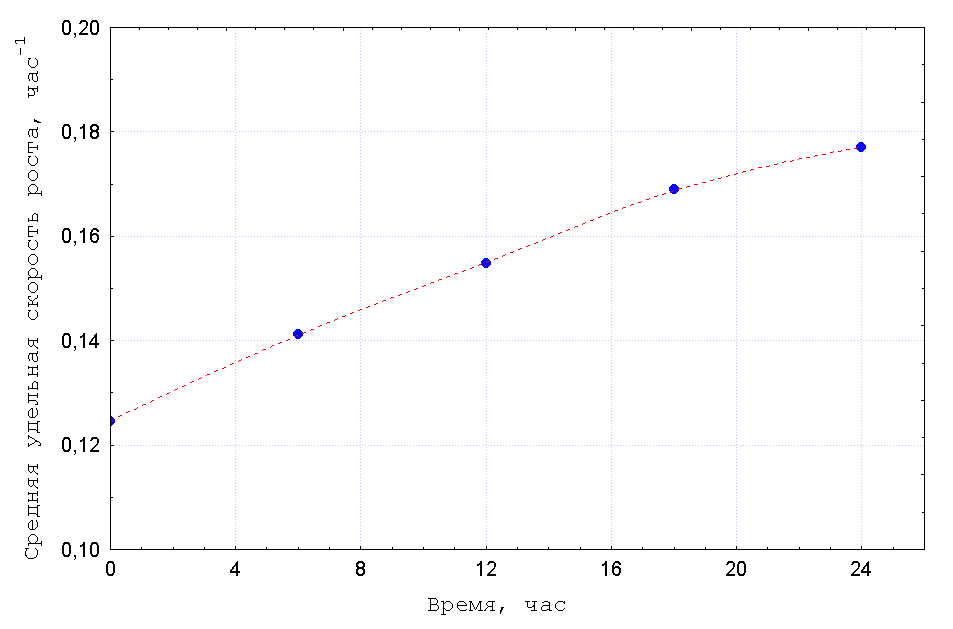
Эволюционная динамика популяций в хемостате является более сложной, т.к.
удельные скорости размножения
![]() являются функциями
времени вследствие нестационарности концентрации лимитирующего
субстрата
являются функциями
времени вследствие нестационарности концентрации лимитирующего
субстрата ![]() . В данном случае в качестве измеряемой (заданной)
величины выберем среднюю по популяции и по времени удельную скорость
роста
. В данном случае в качестве измеряемой (заданной)
величины выберем среднюю по популяции и по времени удельную скорость
роста
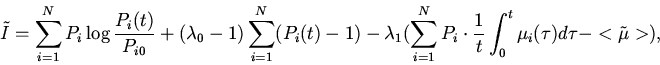
Из (20) и (21) непосредственно получается явное выражение
для производства информации в ходе эволюции хемостатной популяции
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Sunday, 07-Oct-2001 14:05:34 NOVST
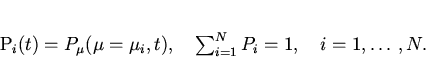
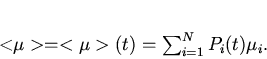
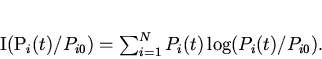
![\begin{displaymath}
\begin{array}{l}
I(P_i(t)/P_{i0})\Rightarrow \min,\\
\delta [I(P_i(t)/P_{i0})]=0
\end{array}
\end{displaymath}](img27.gif)