Моделирование регуляторных механизмов сердечно-сосудистой системы
1
Хидирова М.Б. Национальный Университет Узбекистана, Ташкент
Аннотация:
На примере моделирования простейшим функциональным уравнением
демонстрируется важность учета пространственной разделенности
процессов возникновения и распространения возбуждения по сердечной
ткани. Показана возможность математического моделирования
последовательной активации мышечных клеток сердечной ткани путем
привлечения класса дифференциально-разностных уравнений. Они позволяют
учитывать временные взаимоотношения в системе сердечной регуляции.
Вследствие сложности построенной системы дифференциально-разностных
уравнений предлагаются, для модельных исследований, способы построения
их модельных систем. В работе приведены результаты качественного
анализа разработанных уравнений возбуждения сердечной ткани и
применения их для математического и компьютерного моделирования
регуляторных механизмов сердечно-сосудистой системы.
On the example of modeling using the most simplest functional
equation, the importance of account the spatial divided processes of
excitement origin and propagation on cardiac tissues is demonstrated.
The possibility of mathematical modeling of consequent activation of
cardiac tissue muscle cells by means of differential-delay equations
is shown. This equations allow to take into account the temporary
relations in the system of cardiac regulation. Due to complexity of
constructed system of differential-delay equations, the methods for
construction of their model systems are offered for model studies. In
this work the results of the qualitative analysis of developed
equations of cardiac tissue excitement and using this equations for
mathematical and computer modeling of cardiovascular system regulatory
mechanisms are given.
На фоне неослабевающей остроты сердечно-сосудистых
заболеваний актуально всемерное развитие количественных методов
исследования механизмов кровообращения. Становится очевидным, что
успешное решение данной проблемы возможно только при применении
методов математического и компьютерного моделирования механизмов
кровообращения с учетом закономерностей возбуждения сердечной ткани.
Результаты экспериментальных и модельных исследований
[1,4,5,7]
показывают сложное пространственно-временное разнообразие процессов
возбуждения сердечной ткани. В данной работе рассматриваются возможные
простейшие уравнения возбуждения сердечной ткани, в которых
учитывается пространственная разделенность процессов возникновения и
распространения возбуждения.
Характерными проявлениями процесса возбуждения сердечной ткани
являются электрические изменения.
Активное (возбужденное) мышечное волокно становится отрицательно
заряженным по отношению к неактивному (не возбужденному) мышечному
волокну. Таким образом, возникает разность потенциалов всегда в тех
случаях, когда в процессе сердечной активности часть мышечных волокон
сердца возбуждена, а часть волокон не возбуждена
[8]. Пусть общая
величина разности потенциалов - некоторая функция от времени.
Давление крови, выбрасываемой сердцем в момент времени , в основном
зависит от количества возбужденных клеток в желудочках и в
предсердиях. Следовательно существует функциональная зависимость
между давлением крови (обозначим его через ), выбрасываемой сердцем, и
. Если еще учесть, что давление крови, выбрасываемой желудочками в
момент времени , зависит не только от того, сколько клеток сердца в
этот момент возбуждено, но и от величины давления крови в предыдущие
моменты времени (от начала систолы желудочков), и от величины давления
крови, выбрасываемой предсердиями в желудочки во время систолы, то в
общем случае зависимость между и является очень сложной
функцией и требует применения уравнений, позволяющих учитывать
пространственную разделенность процессов в сердечной деятельности.
Рассмотрим зависимость вида:
(1)
где - непрерывные функции, выражающие радиальную
упругость предсердий и желудочков. Легко заметить, что если нам
известен вид на и - на начальном отрезке
, то этого
достаточно, чтобы найти на при заданных
.
Для анализа простейших особенностей решений (1) найдем вид
при
очень сильных допущениях относительно и , а именно пусть
и - очень малое положительное число. Обозначая
приведем уравнение (1) к виду
(2)
Будем считать, что
. Тогда
или приближенно уравнение (2) с учетом начальных условий запишем в
виде
Теперь исследуем поведение при различных предположениях
относительно .
1. Пусть
. Тогда
Так как очень мало, то имеем колебания с очень малыми
амплитудами. Следовательно могут быть такие электрические изменения на
поверхности сердца, при которых давление потока крови отсутствует, т. е.
могут быть такие "возбуждающие" импульсы, которые не в состоянии
возбудить клетки сердца. Это означает, что существует так называемое
пороговое значение импульса для клеток сердца. Если сила импульса ниже,
чем это пороговое значение, то клетки сердца не могут выполнить
сократительную функцию. Этот факт в медицине известен
[8]. Если даже
сердце остановится, то достаточно долгое время можно наблюдать
ритмические электрические изменения на поверхности сердца.
2. Пусть
. Имеем
Это - колебания около с конечной амплитудой.
Так как , то все
время какая-то часть поверхности желудочка сокращается. Это
способствует непрерывному току крови.
3. Пусть
,
; .
Тогда
Обозначим через члены, содержащие как коэффициент;
- малые колебания. Получим
Здесь мы имеем затухающие колебания.
4. В прикладном моделировании сердечной активности интересным
является случай
(3)
очень близкий к реальной функции артериального давления [8].
Как показали расчеты на ПЭВМ, в данном случае в зависимости от
конкретных значений параметров могут наблюдаться различные, близкие к
реальным, режимы процесса возбуждения сердечной ткани.
Рассмотренное количественное описание сердечной активности на
основе уравнения (1) может быть использовано для исследования процесса
возбуждения сердечной ткани в ограниченном круге задач. Оно является
упрощенным, но отражающим существенные стороны активации сердечной
мышцы уравнением. Следует отметить следующие недостатки:
зависимость активности сердечной деятельности от давления в артериях,
пренебрежение существованием левого и правого отделов сердца,
отсутствие влияния состояния организма на активность сердца.
В работах [2,11,12] изложены результаты предпринятых попыток
моделирования сердечной активности на основе последовательного
количественного описания возбуждения мышечных волокон сердечной
ткани с учетом вышеуказанных сторон сердечной деятельности. Согласно
данной методике, мышечная система сердца рассматривается как
возбудимая среда с водителем ритма (пейсмекером) в синусовом узле. По
мере распространения волн активности по поверхности сердца происходит
функционирование его отделов. Активность пейсмекера регулируется
посредством нейро-гуморальных факторов. Обозначая через:
- величину возбуждения мышечных волокон в момент времени
;
- величину возбуждения мышечных волокон в момент времени
;
. . . . . . . .
- величину возбуждения мышечных волокон в момент времени
,
распространение возбуждения в синусовом узле мышечной ткани сердца
можно описать следующими дифференциально-разностными уравнениями
[11]:
(4)
где - коэффициенты, выражающие скорость возбуждения активности в
слое мышечных волокон; - коэффициенты, выражающие скорость
спада активности в слое мышечных волокон. Следует отметить, что
, а значение
выражает энергетические потери.
Затем волна охватывает оба предсердия, которые имеют общую
мускулатуру:
правое предсердие
(5)
левое предсердие
(6)
где
-
количество возбужденных мышечных волокон
правого и левого предсердий в момент времени . Коэффициенты
того же характера, как и в случае уравнения (4).
Дальнейшее распространение волны возмущения по мышечным
волокнам желудочков выразим через следующие дифференциально-разностные
уравнения:
правый желудочек
(7)
левый желудочек
(8)
где
- количество возбужденных мышечных волокон
правого и левого желудочков. Здесь коэффициенты
-
такого же характера, как и в предыдущих
уравнениях.
Перейдем к составлению уравнений возбуждения мышечных волокон
синусового узла в начале сердечного цикла. Количество и степень
возбуждения этих волокон зависит от энергообеспеченности, от
эффективного функционирования предсердий и желудочков предыдущего
этапа сердечного цикла, от состояния сердца и организма в целом.
Перечисленные параметры в конечном итоге зависят от функционирования
сердечных мышц, на основе которого весь организм обеспечивается необходимыми
ресурсами. Тогда предполагая, что характер возбуждения зависит от
характеристик самой возбудимой среды [4,7,9,13] и
учитывая нейрогуморальное воздействие на клетки синусового узла (со стороны
организма, в зависимости от его состояния, и следовательно от
характера работы сердца), можем написать:
где - функция стимулирования;
- функция ингибирования возбуждения
рассматриваемых клеток.
При исследовании подобных процессов функция ингибирования
обычно берется в виде монотонно убывающей функции от своих
аргументов [9]. Учитывая опыт моделирования замкнутых систем регуляции
[9,10], напишем:
где
- неотрицательные постоянные.
Поскольку функционирование сердца в норме предполагает
синхронную работу желудочков и прекращение активности одного из них
приводит к прекращению и активности сердца в целом, стимулирующую
функцию можно взять в виде однородной функции своих аргументов.
Следовательно, в наиболее упрощенном виде для величины возбуждения
мышечных волокон синусового узла на начальном этапе сердечного цикла
мы можем написать уравнение:
(9)
где - неотрицательная постоянная.
Таким образом, мы получили замкнутую систему из
дифференциально-разностных уравнений нелинейного типа ((4)-(9)).
Значения могут быть определены на основе биологических данных.
При задании начальных условий на соответствующих отрезках времени
решение данной системы уравнений можно получить методом
последовательного интегрирования [9,13].
Для уравнений систем (4)-(8) имеют место нулевые начальные условия,
так как к моменту начала нового цикла мышечные волокна сердца
находятся в покое. Для уравнения (9), выражающего величину
возбуждения мышечных волокон синусового узла на начальном этапе
сердечного цикла, значения коэффициентов и начальные условия
определяются с учетом значений давления крови венозного возврата. В
этом случае первое уравнение (4) представляет собой неоднородное
дифференциальное уравнение вида:
с известной - функцией величины возбуждения мышечных волокон
синусового узла на начальном этапе сердечного цикла и
. Решение данного
уравнения на отрезке длины определяет неоднородную часть второго
уравнения (4) и так далее. Таким путем определяются решения (4)-(8) и
правая часть уравнения (9).
Нелинейность и большое количество рассматриваемых уравнений
требует использования соответствующих редуцированных систем. В работе
[11] подробно изложены основные методы редукции подобных уравнений и
рассмотрен случай предельно редуцированных уравнений возбуждения
сердечной ткани в виде следующего одного нелинейного
дифференциально-разностного уравнения:
(где
)
которое можно написать в следующем безразмерном виде:
(10)
где - величина возбуждения мышечных волокон в момент времени ;
-
параметр, характеризующий среднюю скорость возбуждения волокон
мышечной ткани сердца; - параметр, характеризующий среднюю
скорость подавления возбуждения волокон мышечной ткани сердца; -
параметр, характеризующий среднее время, необходимое для
осуществления обратной связи в системе сердечной деятельности.
Значения параметров и зависят, в основном, от уровня кровоснабжения
сердечной мышцы, а значение - от состояния организма в целом.
Было
показано, что уравнение (10) обладает устойчивыми колебательными
решениями, выражает функционально-активные режимы сердечной
деятельности и может быть применено для количественного и
качественного анализа наиболее общих закономерностей возбуждения
сердечной ткани в целом [2,12].
Оно (включая уравнения Навье-Стокса для
описания движения крови по артериальным сосудам и уравнения активной
и пассивной диффузии для описания транспорта крови по органам и тканям
организма) было использовано при создании минимальной замкнутой
модели кровообращения и программного обеспечения - REGUS [10].
Применение REGUS при анализе механизмов кровообращения в норме, при
возникновении и развитии аритмии и внезапной остановки сердца показало
возможность его использования для количественных исследований
механизмов функционирования сердечно-сосудистой системы. Программа
REGUS была успешно применена при решении задачи оптимизации
хирургического лечения при портальной гипертензии в рамках гранта 40-96
(совместно с сотрудниками НИИ МЗ РУз "Грудная хирургия", под
руководством проф. Адыловой Ф.Т. [3]).
Следует отметить, что при рассмотрении задач анализа возможных
аномалий в предсердиях, желудочках и в пейсмекере может быть
использована (полученная из уравнений (4)-(9)), следующая
редуцированная система уравнений возбуждения сердечной ткани с учетом
средних значений количества возбужденных клеток в пейсмекере,
предсердиях и желудочках:
(11)
где
- переменные, выражающие уровень
возбуждения пейсмекера, предсердий и желудочков;
- положительные постоянные.
Результаты качественных исследований (11) показывают наличие
устойчивого тривиального положения равновесия. Существование
нетривиальных положений равновесия зависит от решений уравнения
(12)
где
Анализ (12) показывает наличие двух положительных корней при
Исследование устойчивости нетривиальных положений равновесия
(11), проведенное на основе качественного и количественного анализа на
ПЭВМ, показывает наличие неустойчивого положения равновесия и области
колебательных решений. Последние могут быть в виде предельных циклов
типа Пуанкаре (нормальная ритмика сердца) и нерегулярных колебаний
(различная форма аритмии). В некоторых случаях, в ходе вычислительных
экспериментов на соответствующей модификации REGUS, происходит
срыв колебаний (эффект "черная дыра"), который выражает внезапную
остановку сердца и резкое падение давления в системе кровообращения.
Детальный качественный анализ (11) с определением основных
закономерностей поведения решения в фазовом пространстве и
построением параметрического портрета является предметом дальнейших
исследований.
Таким образом, в зависимости от решаемых задач в области
количественного анализа сердечной активности, могут быть использованы:
функциональное уравнение (1), дифференциально-разностное уравнение
(10) и система дифференциально-разностных уравнений (11). Приведенные
предварительные результаты их качественного анализа показывают
существование, при определенных условиях, устойчивых стационарных,
периодических решений, нерегулярных колебаний и эффекта "черная дыра".
В случае идентификации условий их существования с конкретными
значениями параметров сердечной деятельности в условиях клиники можно
осуществить моделирование конкретных заболеваний системы
кровообращения [3] и предложить рекомендации для выбора тактики
лечения.
Hidirova M.B. Cybernetic simulation of "heart-vessels" systems. In
proceedings, 10-th International Congress on Cybernetics and Systems.
August, 26-31, 2, Buharest, 1996, 77.
Hidirova M.B., Adilova F.T. Mathematical Modeling of Portal
Hypertension//Proceedings, Ninth World Congress on Medical Informatics,
August 18-22, Korea, Seoul, Edited by B.Cesnik, A.T.McGray,DS, Press.
Amsterdam-Berlin-Oxford-Tokyo-Washington, 1998, 399.
Хидирова М.Б. Программа для гибридного моделирования сердечно-
сосудистой системы (REGUS). Свидетельство № 83. N ED GU 9700014.
Зарегистрировано в Государственном реестре программ для ЭВМ
24.04.1997.
Хидирова М.Б. Об одной замкнутой модели сердечно-сосудистой
системы (ССС): Основные уравнения и результаты вычислительных
экспериментов //Проблемы механики, 5, 1998, 26-30.

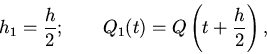
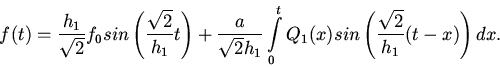
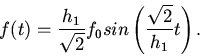
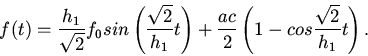
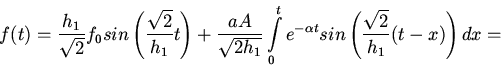
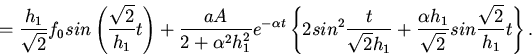
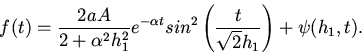
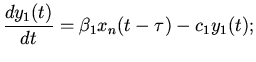
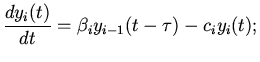
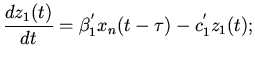
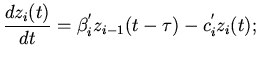
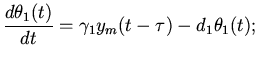
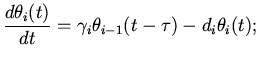
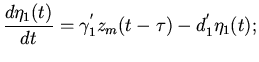
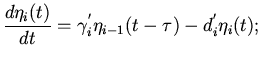
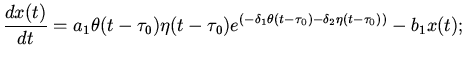
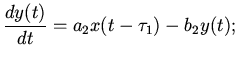
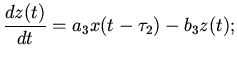
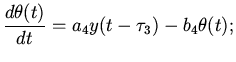
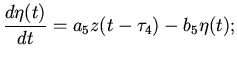
 Ваши комментарии
Ваши комментарии
![[SBRAS]](/gif/s_sigma.gif)