Новосельцев В.Н.
Институт проблем управления РАН, Москва
Яшин А.И.
Max-Planck Institute for Demographic Study, Rostock, Germany
1. Модели истории жизни и распределения ресурсов
Моделирование баланса ресурсов и их роли в формировании истории жизни было начато группой английских исследователей в конце 1980-х гг. [3]. Общая идея состояла в том, что эволюция истории жизни закрепляет в ней такие признаки, которые в данных условиях среды обеспечивают максимальную фишеровскую приспособленность ![]() (максимум среднего по популяции репродуктивного успеха [6,13,25]).
(максимум среднего по популяции репродуктивного успеха [6,13,25]).
Основные достижения на этом пути связаны с работами Ван Ноордвийка и Де Йонга [24], предложившими простую графическую модель генетических корреляций между признаками истории жизни. Кирквуд [10, 11] рассмотрел расход энергии на поддержание сомы ![]() и показал, что существует оптимум
и показал, что существует оптимум ![]() . Было показано, что значение
. Было показано, что значение ![]() меньше гипотетического уровня
меньше гипотетического уровня ![]() "выше которого темп старения предполагается равным нулю". Поэтому появился термин "одноразовая сома" (disposable soma), подчеркивающий аналогию соматических структур тела с такими предметами одноразового пользования, как зубная щетка или бритвенное лезвие, которые выполняют свою функцию,
не будучи слишком надежными [10,20].
"выше которого темп старения предполагается равным нулю". Поэтому появился термин "одноразовая сома" (disposable soma), подчеркивающий аналогию соматических структур тела с такими предметами одноразового пользования, как зубная щетка или бритвенное лезвие, которые выполняют свою функцию,
не будучи слишком надежными [10,20].
Наиболее известная эволюционная модель баланса ресурсов принадлежит Партридж и Бэртону [19]. Они предложили простую модель "оптимальной истории жизни, при которой выживание и репродукция в поздних возрастах приносятся в жертву ранней репродукции и выживанию". В популяция молодых (1) и взрослых (2) самок вероятность дожития до возраста (1) равна ![]() , а от (1) до (2) --
, а от (1) до (2) -- ![]() . Каждая самка в каждом из возрастов производит
. Каждая самка в каждом из возрастов производит ![]() дочерей. Скорость роста популяции
дочерей. Скорость роста популяции ![]() (параметр приспособленности Фишера) связана с
(параметр приспособленности Фишера) связана с ![]() ,
, ![]() и
и ![]() простым уравнением
простым уравнением
![]() .
Комбинация
.
Комбинация ![]() ,
, ![]() и
и ![]() , которая обеспечивает максимальное значение
, которая обеспечивает максимальное значение ![]() , оптимальна. Предполагается, что ресурсы организма ограничены, так что
, оптимальна. Предполагается, что ресурсы организма ограничены, так что ![]() , а
, а ![]() и
и ![]() связаны соотношением
связаны соотношением
![]() . Тем самым в ходе эволюции допускается плейотропическое перераспределение ресурсов. Эти предположения позволили представить процесс эволюционного формирования оптимальных признаков истории жизни в графическом виде: на плоскости с координатами J, A линии уровня r (J, A) пересекаются кривой эволюционного ограничения
. Тем самым в ходе эволюции допускается плейотропическое перераспределение ресурсов. Эти предположения позволили представить процесс эволюционного формирования оптимальных признаков истории жизни в графическом виде: на плоскости с координатами J, A линии уровня r (J, A) пересекаются кривой эволюционного ограничения
![]() . Точка максимума есть точка касания ограничивающей кривой и линии уровня, отвечающей максимуму
. Точка максимума есть точка касания ограничивающей кривой и линии уровня, отвечающей максимуму ![]() .
.
2. Гомеостатическая модель истории жизни
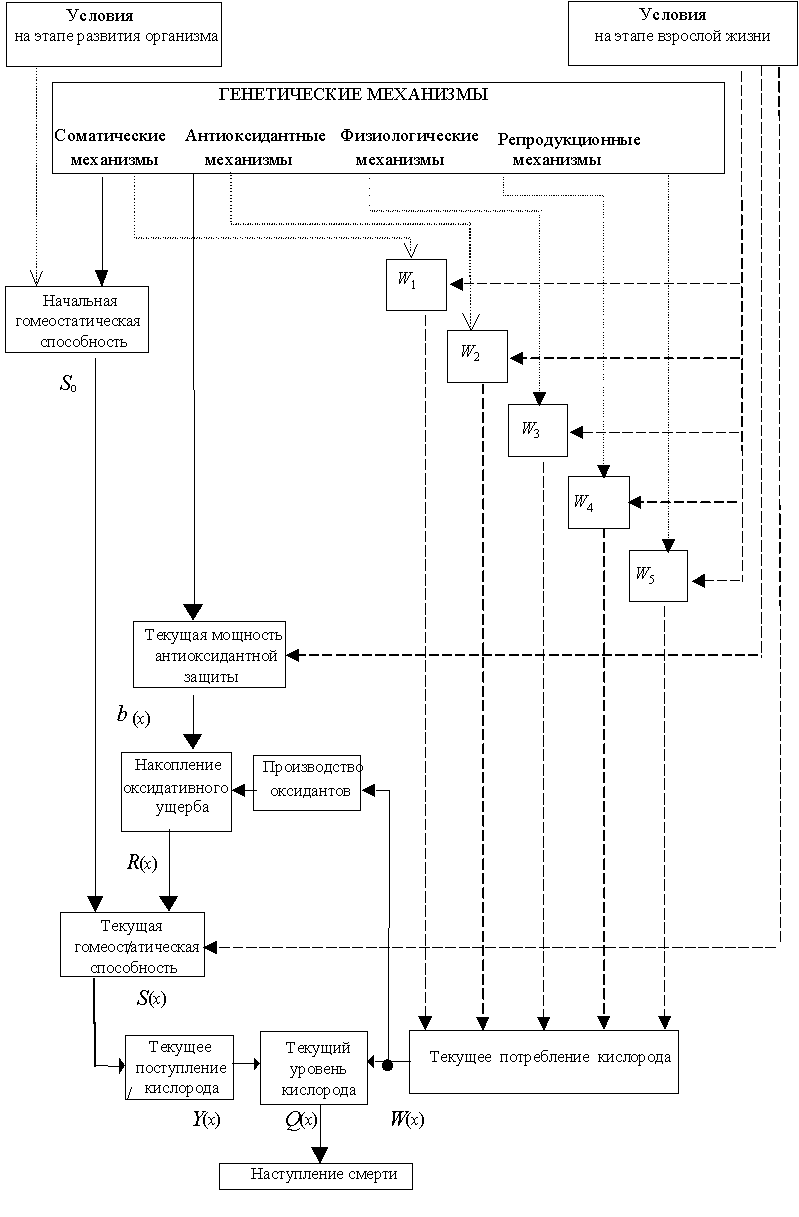
В этой модели история жизни организма представляется как снижение его гомеостатической способности ![]() с возрастом
с возрастом ![]() в связи с репродуктивным функционированием [15-17,30]. Гомеостатическая способность оцениваются с точки зрения энергетического баланса (доставки кислорода). Снижение доставки кислорода с возрастом приводит к падению энергетического ресурса организма. Когда ресурс иссякает, поддержание стационарного состояния делается невозможным и наступает смерть.
в связи с репродуктивным функционированием [15-17,30]. Гомеостатическая способность оцениваются с точки зрения энергетического баланса (доставки кислорода). Снижение доставки кислорода с возрастом приводит к падению энергетического ресурса организма. Когда ресурс иссякает, поддержание стационарного состояния делается невозможным и наступает смерть.
В соответствии с теорией оксидативного стресса [8,29,33], потребление кислорода в возрасте ![]() ,
равное
,
равное ![]() , приводит к образованию оксидативных частиц со скоростью
, приводит к образованию оксидативных частиц со скоростью ![]() . Антиоксидантные системы элиминируют основную часть возникающих частиц, но их малая доля
. Антиоксидантные системы элиминируют основную часть возникающих частиц, но их малая доля ![]() наносит ущерб клеточным структурам. Темп появления повреждений
наносит ущерб клеточным структурам. Темп появления повреждений ![]() в возрасте
в возрасте ![]() пропорционален темпу потребления кислорода
пропорционален темпу потребления кислорода ![]() и клеточной уязвимости
и клеточной уязвимости ![]() :
:
| (1) |
| (2) |
| (3) |
Особенностью гомеостатического подхода является прямое введение в модель смерти от старости
![]() )
)
Общая схема формирования истории жизни животного организма с учетом возможного многообразия его функций приведена на рис. 1. эта модель была использована для воспроизведения классических температурных экспериментов на дрозофиле [17] и для анализа процессов старения у плодовой мушки Ceratitis Capitata [15].
3. Эволюционная оптимальность жизненного цикла у самки
дрозофилы Wayne State
Хотя положение об эволюционной оптимальности истории жизни общеизвестно, прямой демонстрации этого факта для конкретных видов животных не было. Здесь мы рассмотрим генотип самки Wayne State [1], и покажем, что ее история жизни эволюционно оптимальна.
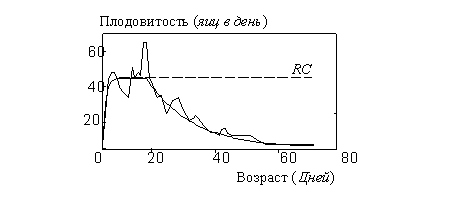
Представим для этого генотипа "репрезентативный организм", описываемый уравнениями (1) - (3) и графиком производства яиц ![]() , показанным на рис. 2 ломаной линией. Зависимость репродуктивных усилий от возраста
, показанным на рис. 2 ломаной линией. Зависимость репродуктивных усилий от возраста ![]() представим как среднеквадратическое приближение экспериментальных данных
представим как среднеквадратическое приближение экспериментальных данных ![]() .
В модели [14] использована зависимость
.
В модели [14] использована зависимость
 |
(4) |
Средний по популяции "репродуктивный успех" организма, ![]() , определяется количеством яиц, произведенных самкой за среднее время жизни (44 дня): согласно (4),
, определяется количеством яиц, произведенных самкой за среднее время жизни (44 дня): согласно (4), ![]() . Зависимость потребления кислорода
. Зависимость потребления кислорода ![]() у дрозофилы имеет вид, показанный на рис. 3.
Поскольку интенсивность производства потомства должна непосредственно отражаться на энерготратах, величину
у дрозофилы имеет вид, показанный на рис. 3.
Поскольку интенсивность производства потомства должна непосредственно отражаться на энерготратах, величину ![]() можно представить как сумму
можно представить как сумму ![]() и
и ![]() (мкл
(мкл![]() /день):
/день):
| (5) |
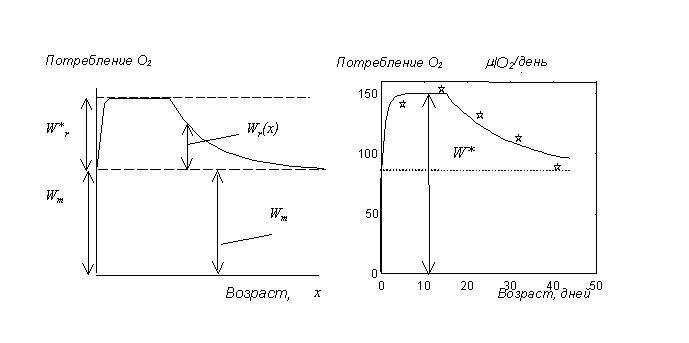 |
Затраты на репродукцию ![]() пропорциональны производству яиц,
пропорциональны производству яиц,
![]() ,
где
,
где ![]() мкл
мкл![]() /яйцо. Максимальное потребление энергии на репродуктивную функцию
/яйцо. Максимальное потребление энергии на репродуктивную функцию
![]() равно
равно
![]() .
Схема моделирования истории жизни у дрозофилы приведена на рис. 4.
.
Схема моделирования истории жизни у дрозофилы приведена на рис. 4.
 |
Для проверки гипотезы эволюционной оптимальности мы используем идею Партридж-Бэртона, подробно рассмотренную выше. Для рассматриваемого генотипа необходимо построить
"фенотипическое множество" -- совокупность теоретически допустимых кривых
![]() . Это множество (линии уровня
. Это множество (линии уровня ![]() для различных комбинаций
для различных комбинаций ![]() и
и ![]() ) получается путем моделирования по схеме рис. 4 генотипа Wayne State (рис. 5).
) получается путем моделирования по схеме рис. 4 генотипа Wayne State (рис. 5).
В соответствии с идеей Партридж-Бэртона, искомый оптимум
![]() лежит в точке касания линии уровня с максимальным репродуктивным успехом,
лежит в точке касания линии уровня с максимальным репродуктивным успехом,
![]() и некоторой кривой, характеризующей "эволюционные ограничения" данного генотипа.
и некоторой кривой, характеризующей "эволюционные ограничения" данного генотипа.
Относительно эволюционных ограничений нами была выдвинута "технологическая гипотеза" [14]. Мы предположили, что генетически запрограммированный уровень репродукции достигается у организма
"в расцвете сил", а старение снижает его (рис. 2). Технологическая гипотеза состоит в том, что на каждую единицу энергии, вложенную в производство потомства, организм должен потратить k единиц энергии во вспомогательных соматических процессах.
Тогда репродуктивную эффективность генотипа, определяемую эволюционными ограничениями, можно описать как КПД этого процесса ![]() . Экспериментальные данные по потреблению кислорода позволяют дать непосредственную оценку величины КПД.
Для генотипа Wayne State (в соответствии с рис. 3):
. Экспериментальные данные по потреблению кислорода позволяют дать непосредственную оценку величины КПД.
Для генотипа Wayne State (в соответствии с рис. 3):
![]() . В декартовых координатах
. В декартовых координатах ![]() )
)
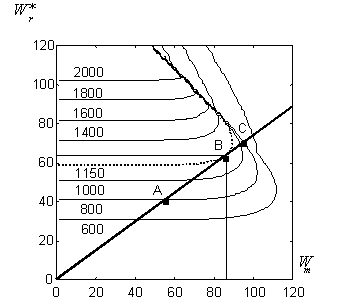 |
Степень эволюционной оптимальности генотипа определяется близостью оценки КПД непосредственно по экспериментальным данным рис. 3 (![]() ) и теоретических построений на рис. 5. В нашем случае ограничительная прямая касается линии уровня, соответствующей экспериментальному репродуктивному успеху самки,
) и теоретических построений на рис. 5. В нашем случае ограничительная прямая касается линии уровня, соответствующей экспериментальному репродуктивному успеху самки, ![]() яиц. Как увеличение, так и уменьшение вклада в репродукцию (при сохранении "технологии" генотипа) приводит к уменьшению репродуктивного успеха (точки
яиц. Как увеличение, так и уменьшение вклада в репродукцию (при сохранении "технологии" генотипа) приводит к уменьшению репродуктивного успеха (точки ![]() и
и ![]() ). Таким образом, гипотеза об эволюционной оптимальности генотипа самки Wayne State вполне соответствует имеющимся теоретическим представлениям и не противоречит экспериментальным данным.
). Таким образом, гипотеза об эволюционной оптимальности генотипа самки Wayne State вполне соответствует имеющимся теоретическим представлениям и не противоречит экспериментальным данным.
4. Прогноз пределов продолжительности жизни у человека
Увеличение энергетической нагрузки ![]() в соответствии с (3) в любом возрасте может привести к истощению ресурса
в соответствии с (3) в любом возрасте может привести к истощению ресурса ![]() )
)![]() в зависимости от возраста, то можно получить наглядную оценку "потенциала поддержания жизни": в соответствии с (1) и (3) экспоненциальное снижение гомеостатической способности S с возрастом приводит к экспоненциальному снижению
в зависимости от возраста, то можно получить наглядную оценку "потенциала поддержания жизни": в соответствии с (1) и (3) экспоненциальное снижение гомеостатической способности S с возрастом приводит к экспоненциальному снижению ![]() .
.
В физиологии человека аналогичная величина известна как максимальное потребление кислорода, МПК, и широко используется при анализе работоспособности организма [2]. Гомеостатический подход открывает возможность использовать накопленные в этой области данные для демографических целей -- прогноза предельной продолжительности жизни.
 |
Причины смерти животных в природе лежат в проксимативной и регуляторной сфере [2,23]. Накопление оксидативного ущерба и истощение кислородного ресурса -- проксимативные процессы, происходящие на клеточном и субклеточном уровне. На них накладываются регуляторные ограничения, отражающие физиологический уровень организации [27,28]. Очевидно, гомеостатический подход в представленном здесь виде ограничен "клеточным" старением и поэтому позволяет определить лишь проксимативный предел продолжительности жизни. Если обозначить минимально необходимый для жизни темп потребления кислорода как мПК, то можно сказать, что естественная смерть от старости (в проксимативном понимании) наступает при МПК < мПК.
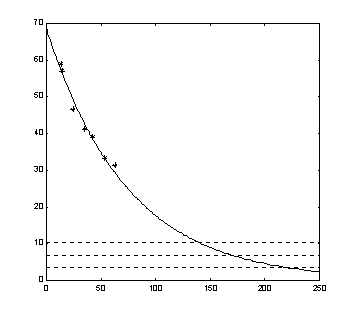
В соответствии с этими представлениями мы привлекаем две группы данных. Первая группа относится к возрастным изменениям максимальных возможностей организма, МПК. Результаты непосредственного изменения МПК с возрастом приводятся в монографии [2] (рис. 6). Вторая группа привлекаемых данных связана с возрастным изменением минимальных потребностей организма, мПК. Принято считать, что величина мПК у человека с возрастом приближается к уровню базального потребления, составляя в пределе около 150
Горизонтальные пунктирные линии на рис. 6 соответствуют различным мПК (100, 200 и 300% от базального уровня человека в 70 лет). Соответствующие оценки проксимативного предела жизни составляют 217, 169 и 138 лет. С учетом представлений работы [12] (превышение базального уровня в 150%), окончательной оценкой проксимативного предела жизни человека является цифра 190 лет.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Friday, 07-Sep-2001 17:05:19 NOVST