Кизилова Н.Н.
Харьковский национальный университет
Distant transport of a liquid in plants and animals takes place in special branching channels -- arterial and vein systems in animals, xylem and phloem -- in plants. Results of quantitative research of parameters of conducting systems in dicotyledonous leaves are presented. There are a number of dependences between diameters, diameters and branching angles of consecutive elements and others, similar to the dependences which have been found out in arterial systems in animals. Rouh' rules and Murray's law are carried out in conducting bands. For all investigated leaves there is a good agreement between the results of measurements and parameters of the optimum pipeline which provides delivering of a liquid with a minimal cost. It is supposed, that possible basis of formation of plant conducting structures is the system of the feedback providing a balance between inflow and consumption of a liquid in a leaf.
В соответствии с подходом, сформулированным в работах А.А.Ляпунова, ``систематизация эмпирических данных в биологии должна проводиться под некоторым единым углом зрения, одинаково существенным для всех разделов биологии'' [14]. Этот подход как нельзя лучше подходит для совместного анализа проводящих систем растений и животных, особенности метаболизма клеток которых и физические основы движения жидкостей по проводящим элементам имеют существенные отличия. В качестве такого единого угла зрения в данной работе предлагается принцип конструирования оптимальных транспортных структур. В общем виде был он был сформулирован в работах Н.Рашевски как принцип оптимальной конструкции [17]. Утверждения об оптимальной организации живых организмов неоднократно высказывались и обсуждались в различных разделах биологии. В соответствии с предостережениями А.А.Ляпунова относительно понимания оптимальности биосистемы [14] в данной работе рассматривается оптимальность проводящей системы относительно ряда критериев, соответствие которым обеспечивает доставку жидкости по системе с минимальными для организма затратами. При моделировании артериальных русел в качестве критериев оптимальности рассматриваются полный объем, площадь поверхности, гидравлическое сопротивление, рассеиваемая мощность, трение на стенках и время доставки крови в конечные элементы русла. Закономерности организации проводящих пучков листьев растений лучше всего соответствуют модели оптимальной ветвящейся системы, обеспечивающей при заданном полном объеме минимальное полное гидравлическое сопротивление (или мощность, которая при фиксированном объемном расходе отличается от гидравлического сопротивления только константой) [12].
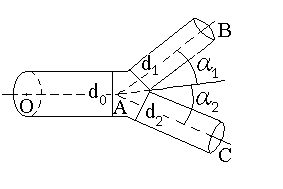
Закономерности ветвления артериальных систем были обобщены В.Ру [18]
и сформулированы в виде набора правил ветвления произвольной артерии
с диаметром ![]() (рис. 1) на две дочерних артерии с диаметрами
(рис. 1) на две дочерних артерии с диаметрами ![]() и
и ![]() и углами ветвления
и углами ветвления ![]() и
и ![]() соответственно:
соответственно:
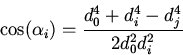 |
(1) |
С.Мюрреем была предложена модель оптимального ветвления, обеспечивающего
минимум полной работы системы ![]() - гидравлической, связанной с
продвижением жидкости по трубкам, и метаболической, связанной c поддержкой
состояния стенок самих сосудов [4]. Для случая ламинарного течения
ньютоновской жидкости по твердой недеформируемой трубке за счет перепада
давления на ее концах (пуазейлевское течение) объемный расход жидкости
- гидравлической, связанной с
продвижением жидкости по трубкам, и метаболической, связанной c поддержкой
состояния стенок самих сосудов [4]. Для случая ламинарного течения
ньютоновской жидкости по твердой недеформируемой трубке за счет перепада
давления на ее концах (пуазейлевское течение) объемный расход жидкости ![]() для оптимальной трубки, обеспечивающей минимум
для оптимальной трубки, обеспечивающей минимум ![]() при заданном объеме,
пропорционален кубу ее диаметра:
при заданном объеме,
пропорционален кубу ее диаметра: ![]() . Если при движении через
ветвление расход сохраняется, а каждый из элементов оптимален в
вышеуказанном смысле, то
. Если при движении через
ветвление расход сохраняется, а каждый из элементов оптимален в
вышеуказанном смысле, то ![]() ,
, ![]() ,
, ![]() . Отсюда
следует закон Мюррея:
. Отсюда
следует закон Мюррея:
который по значению и простоте формулировки часто сравнивают с теоремой Пифагора в геометрии. Многочисленные измерения на препаратах артериальных систем разных органов человека и животных подтвердили справедливость этого соотношения [5,7,9]. Для ветвлений аорты и ряда других систем сосудов, выполняющих не только функцию доставки жидкости, но и ряд иных функций, верен обобщенный закон Мюррея в виде:
| (2) |
Соотношения (1) и (2) описывают основные закономерности строения
артериальных систем, но не позволяют сконструировать русло единственным
образом, поскольку три соотношения (1)-(2) содержат пять параметров
![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() . Для компьютерного
моделирования оптимальных в указанном смысле проводящих систем приходится
вводить дополнительные ограничения, например, значения параметров,
характеризующих асимметрию бифуркаций (
. Для компьютерного
моделирования оптимальных в указанном смысле проводящих систем приходится
вводить дополнительные ограничения, например, значения параметров,
характеризующих асимметрию бифуркаций (![]() ,
, ![]() и
др.) [7], а также задавать правила для определения длин элементов
и
др.) [7], а также задавать правила для определения длин элементов
![]() ,
, ![]() ,
, ![]() каждой бифуркации. Возможно, именно отсутствие жестких связей
между всеми параметрами проводящей системы обеспечивает лабильность живых
организмов, расширяя возможности адаптационных перестроек. Во всех случаях
при измерениях на препаратах артерий обнаруживается значительный разброс
всех вышеперечисленных параметров относительно их оптимальных значений.
При этом значения параметра, осредненные по всему массиву данных, весьма
точно соответствуют оптимальным. Специальные исследования показали, что
в каждой отдельной бифуркации артерий диаметры и углы ветвления могут в
несколько раз отличаться от рассчитанных по (1-2) оптимальных значений,
однако эти изменения согласованы между собой таким образом, что полная
работа
каждой бифуркации. Возможно, именно отсутствие жестких связей
между всеми параметрами проводящей системы обеспечивает лабильность живых
организмов, расширяя возможности адаптационных перестроек. Во всех случаях
при измерениях на препаратах артерий обнаруживается значительный разброс
всех вышеперечисленных параметров относительно их оптимальных значений.
При этом значения параметра, осредненные по всему массиву данных, весьма
точно соответствуют оптимальным. Специальные исследования показали, что
в каждой отдельной бифуркации артерий диаметры и углы ветвления могут в
несколько раз отличаться от рассчитанных по (1-2) оптимальных значений,
однако эти изменения согласованы между собой таким образом, что полная
работа ![]() для каждой из бифуркаций отличается от предсказываемой моделью
оптимальной величины незначительно, например, для коронарных артерий
человека - не более чем на
для каждой из бифуркаций отличается от предсказываемой моделью
оптимальной величины незначительно, например, для коронарных артерий
человека - не более чем на ![]() [9].
[9].
Организация оптимальной транспортной системы в растущем органе невозможна
без участия управления, основанного на системе обратных связей. Принципы
организации оптимальных артериальных русел получили возможность обоснования
после открытия механочувствительности сосудистого эндотелия, клетки
которого избирательно реагируют на изменение напряжения трения ![]() со
стороны потока крови. При пуазейлевском течении напряжение на стенке
трубки
со
стороны потока крови. При пуазейлевском течении напряжение на стенке
трубки
![]() , поэтому для сосуда, в котором может
поддерживаться
, поэтому для сосуда, в котором может
поддерживаться ![]() , выполняется соотношение
, выполняется соотношение ![]() , что
соответствует модели Мюррея. Таким образом, в сосудах животных существуют
системы управления на клеточном уровне, которые могут обеспечить
соответствие параметров русла оптимальным, поддерживая значение
, что
соответствует модели Мюррея. Таким образом, в сосудах животных существуют
системы управления на клеточном уровне, которые могут обеспечить
соответствие параметров русла оптимальным, поддерживая значение ![]() на
стенке в определенном диапазоне [3]. При этом отклонение угла
ветвления от оптимального может приводить к значительному увеличению
вязких напряжений на внешней или внутренней стенке сосуда и вызывать
комплексную реакцию сосудистого эндотелия [3,6], следствием которой
станет регуляция угла ветвления. Подобные перестройки происходят как в
молодом развивающемся организме, так и во взрослом при адаптации к
изменившимся условиям (например, при появлении новообразований) или при
пересадке тканей. Диаметр артерий также регулируется локально посредством
системы обратных связей, обеспечивающих постоянство
на
стенке в определенном диапазоне [3]. При этом отклонение угла
ветвления от оптимального может приводить к значительному увеличению
вязких напряжений на внешней или внутренней стенке сосуда и вызывать
комплексную реакцию сосудистого эндотелия [3,6], следствием которой
станет регуляция угла ветвления. Подобные перестройки происходят как в
молодом развивающемся организме, так и во взрослом при адаптации к
изменившимся условиям (например, при появлении новообразований) или при
пересадке тканей. Диаметр артерий также регулируется локально посредством
системы обратных связей, обеспечивающих постоянство ![]() на стенке
сосуда, причем после удаления сосудистого эндотелия регуляция исчезает [3].
на стенке
сосуда, причем после удаления сосудистого эндотелия регуляция исчезает [3].
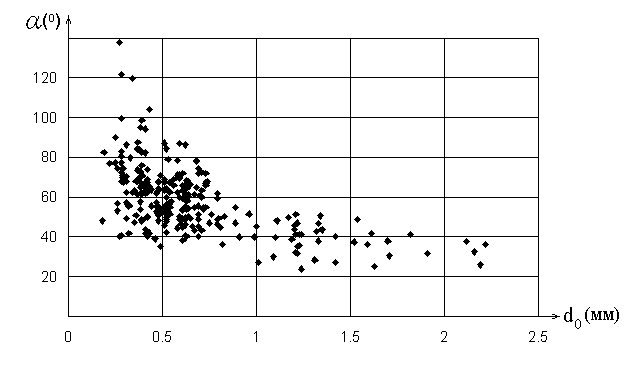
Результаты измерения параметров ветвления элементов проводящей системы
листьев двудольных показали, что существует зависимость полного угла
ветвления
![]() от диаметра
от диаметра ![]() основного
проводящего элемента, причем вид этой зависимости имеет сходный характер
у всех исследованных типов листьев и не зависит от возраста и размера
листа (рис.2). Аналогичные распределения
основного
проводящего элемента, причем вид этой зависимости имеет сходный характер
у всех исследованных типов листьев и не зависит от возраста и размера
листа (рис.2). Аналогичные распределения
![]() для артериальных
русел представлены в [2,15,16] и имеют сходный характер. Исходя из
вида зависимости
для артериальных
русел представлены в [2,15,16] и имеют сходный характер. Исходя из
вида зависимости
![]() , можно выделить подсистемы крупных
(I тип) и мелких (II тип) жилок, закономерности ветвления которых
существенно отличаются. Угол ветвления жилок I типа постоянный или
линейно растет с уменьшением диаметра основного элемента. Для II типа
допустимы значительные вариации угла ветвления (
, можно выделить подсистемы крупных
(I тип) и мелких (II тип) жилок, закономерности ветвления которых
существенно отличаются. Угол ветвления жилок I типа постоянный или
линейно растет с уменьшением диаметра основного элемента. Для II типа
допустимы значительные вариации угла ветвления (![]() ). Можно
сопоставить I и II типы жилок листьев с подсистемами ``распределительных''
(distributing vessels) и ``доставочных'' (delivering vessels) артериальных
сосудов [10], которые отличаются по функциям и имеют аналогичные
различия зависимостей
). Можно
сопоставить I и II типы жилок листьев с подсистемами ``распределительных''
(distributing vessels) и ``доставочных'' (delivering vessels) артериальных
сосудов [10], которые отличаются по функциям и имеют аналогичные
различия зависимостей
![]() .
.
В бифуркациях жилок листьев, как и в сосудистых системах животных, соблюдаются правила Ру:
более крупный дочерний элемент имеет меньший угол ветвления, малые элементы
последних порядков ветвления отходят практически под прямым углом к
основному, дочерние сосуды с равными диаметрами имеют близкие по величине
углы ветвления. Для всех исследованных видов выявлена также зависимость
коэффициента ветвления ![]() (
(
![]() ) от диаметра
основного элемента
) от диаметра
основного элемента ![]() . Выделенные выше подсемейства жилок отличаются
по значениям коэффициента ветвления: элементы
. Выделенные выше подсемейства жилок отличаются
по значениям коэффициента ветвления: элементы ![]() типа имеют
типа имеют ![]() , а
у элементов II типа
, а
у элементов II типа
![]() .
.
Для крупных проводящих элементов листьев выполняется закон Мюррея (2)
с
![]() . Для подсистемы
. Для подсистемы ![]() типа значения
типа значения
![]() лежат в диапазоне
лежат в диапазоне
![]() , а средние для каждого листа
значения
, а средние для каждого листа
значения
![]() . Сравнение измеренных углов ветвления
. Сравнение измеренных углов ветвления ![]() и оптимальных углов
и оптимальных углов ![]() , рассчитанных по (1), показало их
хорошее согласование. Средние значения безразмерной величины
, рассчитанных по (1), показало их
хорошее согласование. Средние значения безразмерной величины
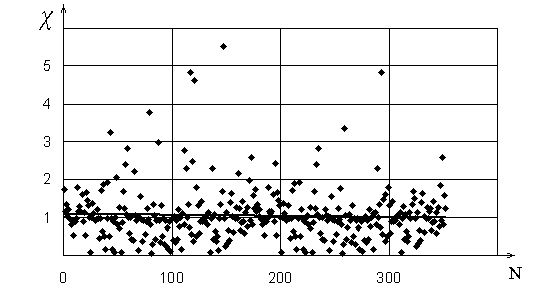
![]() у всех исследованных листьев близки к единице (
у всех исследованных листьев близки к единице (
![]() ,
,
![]() ). Характерное распределение значений
). Характерное распределение значений ![]() приведено
на рис. 3. Лучшее соответствие угла ветвления оптимальному отмечено при
приведено
на рис. 3. Лучшее соответствие угла ветвления оптимальному отмечено при
![]() , что соответствует большинству ветвлений мелких жилок II
типа.
, что соответствует большинству ветвлений мелких жилок II
типа.
 |
Таким образом, полученные закономерности позволяют утверждать, что в
конструкции проводящей системы листа в виде зависимостей между диаметрами
и углами ветвления реализованы принципы, соответствующие модели оптимального
трубопровода, который среди всех трубопроводов с тем же полным объемом
обеспечивает перемещение растительного сока с минимальными затратами.
Существует ли в листе и на каком уровне система регуляции, которая может
отслеживать локальные отклонения параметров проводящего пучка от оптимальных
и изменять их, пока остается неясным. Механорецепторные системы,
подобные таковым в артериях, пока не обнаружены. Помимо этого, сосуды
проводящей системы листа представляют собой полые трубки, состоящие из
клеточных стенок, лишенных живого клеточного содержимого, а значит они не могут отслеживать и
корректировать диаметры и углы ветвления. В развивающемся зачатке листа на стадии
закладки проводящих элементов клетки последних являются живыми, делятся и
могли бы участвовать в локальной регуляции, однако на этой стадии они не
выполняют еще проводящей функции. Возможно, процесс ``оптимизации'' проводящей
системы листа может основываться на поддержании в развивающемся зачатке
листа баланса между притоком жидкости по проводящей системе и ее
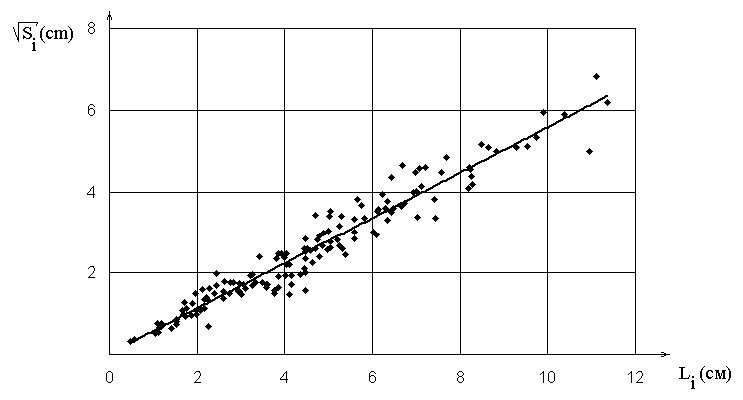
потреблением клетками. Наши измерения показали (рис. 4), что существует
зависимость между длиной проводящего элемента ![]() и площадью
и площадью
![]() той части листовой пластинки, транспорт жидкости к которой
производится по этому элементу:
той части листовой пластинки, транспорт жидкости к которой
производится по этому элементу:
![]() . Качественно
полученная зависимость может быть сформулирована в следующем виде:
более развитая жилка обеспечивает жидкостью большую площадь листа.
Особенно явно эта зависимость отслеживается на асимметричных листьях с
неодинаково развитыми боковыми жилками. При этом при значительных
вариациях формы областей (от полукруга до невыпуклых криволинейных
многоугольников) для большинства исследованных видов листьев значения
. Качественно
полученная зависимость может быть сформулирована в следующем виде:
более развитая жилка обеспечивает жидкостью большую площадь листа.
Особенно явно эта зависимость отслеживается на асимметричных листьях с
неодинаково развитыми боковыми жилками. При этом при значительных
вариациях формы областей (от полукруга до невыпуклых криволинейных
многоугольников) для большинства исследованных видов листьев значения
![]() лежат в узком диапазоне:
лежат в узком диапазоне:
![]() , поэтому
зависимость не связана с простым геометрическим подобием.
, поэтому
зависимость не связана с простым геометрическим подобием.
Клетки-потребители принимают непосредственное участие в перемещении жидкости по проводящей системе (верхний двигатель) и фактором, который может ограничивать объемный ток по проводящей системе, является площадь просвета элементов первых генераций. От расположения листа на побеге (то есть от числа потребителей, расположенных ниже по отношению к восходящему току растительного сока) зависят некоторые параметры жилкования, например, количество мелких жилок на единицу площади листа [12]. В зависимости от количества поступающей в растение жидкости из зачатка может сформироваться крупный или мелкий лист (например, при обилии влаги и при засухе). Многие факты говорят в пользу высказанного предположения, однако на сегодняшний день количественные соотношения между притоком и потреблением жидкости в листе исследованы недостаточно. Другой возможный механизм отслеживания оптимальных локальных параметров транспортной системы может быть связан с регуляцией роста и развития проводящей системы посредством фитогормонов, ряд которых синтезируется в растущем листе и стимулирует деления клеток листа и дифференцировку проводящих пучков.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Wednesday, 05-Sep-2001 21:38:46 NOVST