
Титлянова А.А.
Институт почвоведения и агрохимии СО РАН, Новосибирск
Разрабатывая приложение системного подхода к биологическому круговороту, БК, А.А.Ляпунов выделил три режима БК: стационарный, периодический и переходной.
Для характеристики режимов БК углерода используются следующие параметры:
![]() - запасы вещества в компонентах экосистемы;
- запасы вещества в компонентах экосистемы; ![]() -
интенсивности обменных процессов;
-
интенсивности обменных процессов; ![]() - входной,
- входной, ![]() - выходной потоки в и из экосистемы;
- выходной потоки в и из экосистемы; ![]() - чистая
первичная продукция, т.е. то количество органического вещества, которое
формируется всеми автотрофами экосистемы на единице площади за единицу
времени;
- чистая
первичная продукция, т.е. то количество органического вещества, которое
формируется всеми автотрофами экосистемы на единице площади за единицу
времени; ![]() - гетеротрофное дыхание, т.е. то количество органического
вещества, которое минерализуется всеми гетеротрофами на единице площади за
единицу времени.
- гетеротрофное дыхание, т.е. то количество органического
вещества, которое минерализуется всеми гетеротрофами на единице площади за
единицу времени.
Стационарный режим характеризуется тем, что при неменяющихся
условиях интенсивности всех процессов и запасы вещества во всех компонентах
остаются постоянными, вход вещества в экосистему равен выходу из нее, чистая
первичная продукция равна гетеротрофному дыханию: ![]() ,
,
![]() ,
,
![]() ,
, ![]() .
Стационарный режим осуществляется только в закрытых
искусственных системах.
.
Стационарный режим осуществляется только в закрытых
искусственных системах.
Периодический режим отличается тем, что изменение всех параметров
происходит циклически под воздействием периодически меняющихся факторов
внешней среды. В периодическом режиме в среднем по периоду в многолетнем
разрезе все параметры остаются постоянными. Периодический режим
отождествляется со стационарным, так как за некоторое количество лет при
данном типе функционирования выполняются равенства: ![]() ,
,
![]() ,
,
![]() ,
, ![]()
В основе периодического режима биологического круговорота лежат регулярные циклы, связанные с периодическими изменениями внешней среды, онтогенезом растений и колебательными процессами, которые характерны для сложных сообществ почвенной биоты. Кроме регулярных - суточных, сезонных и многолетних циклов, в которые вписана динамика биологического круговорота, в природе постоянно наблюдаются флюктуации всех его параметров. Флюктуации носят незакономерный характер и, чаще всего, вызываются не одной, а сочетанием нескольких причин. Сочетания могут меняться от года к году, а вместе с ними будут меняться вид и величина флюктуаций.
Базовой составляющей многолетней динамики параметров биологического круговорота являются сезонные циклы, регулируемые изменением гидротермических условий в течение года.
В умеренной зоне, где основной фактор, определяющий смену сезонов, - тепло, непосредственный ответ экосистемы на весеннее повышение температуры заключается в размножении и развитии всех живых организмов.
В связи с началом вегетации растений начинает активно функционировать продукционная часть углеродного цикла: фотосинтез - дыхание - продукция. В экосистеме образуются новые порции органического вещества, создается надземная и подземная фитомасса. Одновременно с продукционным процессом включается процесс отмирания отдельных органов и целых растений. Отмирание фитомассы приводит к образованию мортмассы, т.е. к формированию новых пищевых ресурсов для гетеротрофов.
Гетеротрофы, активность которых с повышением температуры весной возрастает, начинают ускоренно перерабатывать как старую, так и вновь образуемую мортмассу, обогащенную легко минерализуемыми соединениями. Деструкционные процессы интенсифицируются.
Падение температуры ниже нуля приводит к почти полному прекращению метаболизма автотрофов и значительному замедлению метаболизма гетеротрофов. Биологический круговорот в экосистемах, где почвы промерзают, замирает до следующей весны. В экосистемах с незамерзающими почвами гетеротрофное звено, хотя и с низкой интенсивностью, продолжает зимой функционировать. Повышение температуры весной вновь включает автотрофные и резко стимулирует гетеротрофные обменные процессы.
Сезонный (или годичный) цикл биологического круговорота отражается в динамике запасов всех компонентов экосистемы и в закономерном изменении интенсивностей всех обменных процессов.
В периодическом режиме функционируют климаксовые, субклимаксовые экосистемы, часть полуприродных экосистем (пастбища) и длительно существующие антропогенные трансформаты.
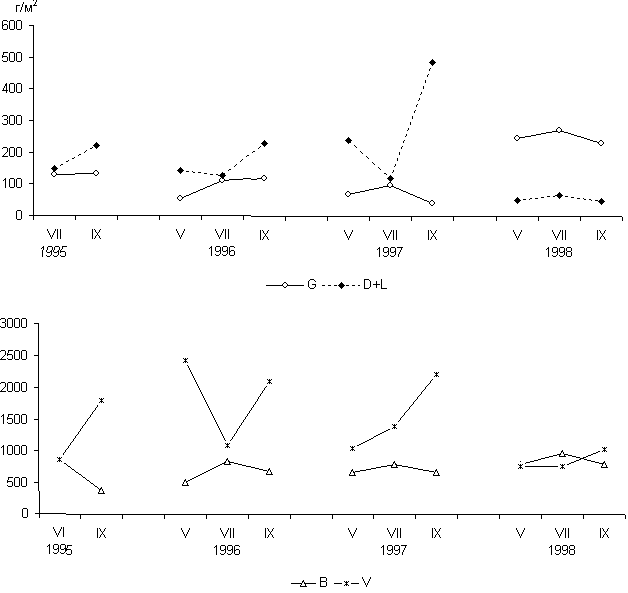
Пример периодического режима - изменение запасов растительного вещества в течение трех лет в пастбищной экосистеме сухой степи. Сухая степь расположена в Туве, в Убсунурской котловине. Режим использования пастбища постоянен - зимний умеренный выпас скота.
Запасы растительного вещества включают: зеленую фитомассу ![]() ; надземную
мертвую фитомассу
; надземную
мертвую фитомассу ![]() , где
, где ![]() - стоящие на корню отмершие растения и
- стоящие на корню отмершие растения и ![]() -
подстилка; подземную живую фитомассу (корни и корневища) в слое почвы 0-20
см.
-
подстилка; подземную живую фитомассу (корни и корневища) в слое почвы 0-20
см. ![]() ; подземную мертвую фитомассу в том же слое почвы
; подземную мертвую фитомассу в том же слое почвы ![]() (рис. 1). Запасы
всех компонентов изменяются периодически. Живая фитомасса (
(рис. 1). Запасы
всех компонентов изменяются периодически. Живая фитомасса (![]() и
и ![]() ) достигает
максимума обычно в середине лета и уменьшается в связи с отмиранием к осени.
Мертвая фитомасса (
) достигает
максимума обычно в середине лета и уменьшается в связи с отмиранием к осени.
Мертвая фитомасса (![]() и
и ![]() ) накапливается к осени и уменьшается в
позднеосенний-ранневесенний периоды, подвергаясь минерализации. В отдельные
годы минерализация происходит активно в летние месяцы после выпадения
дождей. Так, минимум
) накапливается к осени и уменьшается в
позднеосенний-ранневесенний периоды, подвергаясь минерализации. В отдельные
годы минерализация происходит активно в летние месяцы после выпадения
дождей. Так, минимум ![]() отмечался в июле 1997 г., а минимум
отмечался в июле 1997 г., а минимум ![]() в июле 1996 г.
Подобные флюктуации характерны для травяных экосистем.
в июле 1996 г.
Подобные флюктуации характерны для травяных экосистем.
При сохранении периодического режима в разные сезоны ![]() может быть больше
может быть больше
![]() , и тогда общий запас растительного вещества возрастает, или меньше
, и тогда общий запас растительного вещества возрастает, или меньше
![]() , и тогда общий запас уменьшается. В среднем за пятилетний период при
периодическом режиме
, и тогда общий запас уменьшается. В среднем за пятилетний период при
периодическом режиме ![]() и условия стационарности сохраняются, что было
показано на примере луговой степи (табл. 1).
и условия стационарности сохраняются, что было
показано на примере луговой степи (табл. 1).
| Показатель, гС/м |
1972 | 1973 | 1981 | 1982 | 1983 | |
| 624 | 554 | 774 | 1337 | 1572 | 4861 | |
| 444 | 662 | 1274 | 1406 | 1081 | 4867 | |
|
|
+180 | -108 | -500 | -69 | +491 | -6 |
| 29 | -19 | -65 | -5 | 31 | -0,1 |
Переходной режим устанавливается в тех случаях, когда экосистема переходит от одного периодического режима к другому. При одних переходах экосистема может терять вещество и энергию, при других - накапливать.
Так же как и периодический, переходной режим на многолетней шкале складывается из годовых циклов, которые флюктуируют от года к году. Однако, если при периодическом режиме все параметры биологического круговорота флюктуируют вокруг средних величин, то при переходном режиме, колеблясь, они последовательно уменьшаются или увеличиваются до выхода на новый стационарный уровень.
В переходном режиме функционируют все экосистемы, находящиеся на любой стадии любого вида сукцессии, основная часть полуприродных экосистем и большинство агроэкосистем. Следовательно, большая часть биогеоценотического покрова Земли существуют в том или ином переходном режиме.
В переходном режиме с потерями запасы вещества убывают со временем,
вход меньше выхода, читая первичная продукция не равна гетеротрофному
дыханию. При данном режиме выполняются следующие условия:
![]() ,
,
![]() ,
,
![]() ,
,
![]() , где
, где ![]() и
и ![]() - запасы вещества в экосистеме на момент
времени
- запасы вещества в экосистеме на момент
времени ![]() и
и ![]() .
.
Пример переходного режима с потерями - трансформация степной экосистемы в агроценоз. Работы проводились в
Казахстане, в экосистеме настоящей степи и в агроценозе пшеницы, который
возделывался в течение 8 лет после распашки целинной степи по схеме: пар (1
год) ![]() пшеница (3 года)
пшеница (3 года) ![]() пар (1 год)
пар (1 год) ![]() пшеница (3 года).
Оценивались запасы живой фитомассы (
пшеница (3 года).
Оценивались запасы живой фитомассы (![]() ), мертвой фитомассы (
), мертвой фитомассы (![]() ),
почвенного органического вещества в слое почвы 0-20 см (
),
почвенного органического вещества в слое почвы 0-20 см (![]() ), чистая
первичная продукция (
), чистая
первичная продукция (![]() ) и вынос урожая
) и вынос урожая ![]() . Все
данные приводятся в углероде за период 8 лет (табл. 2).
. Все
данные приводятся в углероде за период 8 лет (табл. 2).
| г С/м |
г С/м |
|||||||
| Экосистема | D+L+V |
|
||||||
| Степь | 740 | 675 | 5570 | 8210 | 0 | 8300 | -90 | -1,1 |
| Агроценоз | 310 | 180 | 4710 | 2985 | 420 | 4350 | -1785 | -59,8 |
Степь функционирует в периодическом режиме и отклонение от баланса
составляет всего лишь 1% от ![]() .
.
После распашки целины система входит в переходной режим. Многовидовой
высокопродуктивный степной фитоценоз одномоментно замещается вначале паром
(вспаханное незасеянное поле), затем монокультурой пшеницы, чья чистая
первичная продукция в два раза ниже, чем в степи. С учетом паровых полей (2
года), когда продукция не создается, величина ![]() агроценоза составляет лишь
36% от
агроценоза составляет лишь
36% от ![]() степи. Часть продукции выносится с урожаем. Поступление
растительных остатков, из которых ежегодно образуется некоторое количество
почвенного органического вещества (
степи. Часть продукции выносится с урожаем. Поступление
растительных остатков, из которых ежегодно образуется некоторое количество
почвенного органического вещества (![]() ), в агроценозе резко снижается. В то
же время почва с ее биохимическими свойствами и биотой ``переходит по
наследству'' из степной экосистемы в агроценоз. Какое-то время почва
сохраняет прежнюю потенцию к минерализации
), в агроценозе резко снижается. В то
же время почва с ее биохимическими свойствами и биотой ``переходит по
наследству'' из степной экосистемы в агроценоз. Какое-то время почва
сохраняет прежнюю потенцию к минерализации ![]() . Количество ежегодно
образующегося почвенного органического вещества становится меньше его
ежегодной минерализации. Вынос
. Количество ежегодно
образующегося почвенного органического вещества становится меньше его
ежегодной минерализации. Вынос ![]() с урожаем и потери углерода в виде
с урожаем и потери углерода в виде
![]() при гетеротрофном дыхании,
при гетеротрофном дыхании, ![]() , в сумме превосходят
, в сумме превосходят ![]() (см.табл. 2). Возникает нарушение бюджета углерода, запасы
(см.табл. 2). Возникает нарушение бюджета углерода, запасы ![]() ,
, ![]() и
и ![]() уменьшаются.
уменьшаются.
В ``молодом'' агроценозе осуществляется переходной режим с потерями. С
течением времени при постоянстве ![]() и
и ![]() в
агроценозе установится новое стационарное состояние. Модельные расчеты
показали, что стационарный уровень наступает через 200 лет использования
почв, содержание
в
агроценозе установится новое стационарное состояние. Модельные расчеты
показали, что стационарный уровень наступает через 200 лет использования
почв, содержание ![]() падает до 3680 г С/м
падает до 3680 г С/м![]() и его потери
составляют 36% от исходного запаса (Титлянова и др., 1982).
и его потери
составляют 36% от исходного запаса (Титлянова и др., 1982).
В переходном режиме с накоплением запасы вещества в экосистеме
нарастают, вход больше выхода и (или) чистая первичная продукция больше
гетеротрофного дыхания. Выполняются следующие соотношения:
![]() ,
,
![]() ,
,
![]() ,
, ![]() и (или)
и (или)
![]() .
.
Пример переходного режима с накоплением - сенокосные луга при внесении удобрений. Исследования проводились на
косимых суходольных лугах Чехословакии (Ulehlova et al., 1981; Tesarova,
1983). В опыте было три варианта: луг без удобрений, N-0, луг с внесением
100 кг азота/га, N-1, луг с внесением 200 кг азота/га, ![]() . Оценивались
следующие параметры: запасы углерода надземной фитомассы (G+D+L) и суммы
живых и мертвых подземных органов
. Оценивались
следующие параметры: запасы углерода надземной фитомассы (G+D+L) и суммы
живых и мертвых подземных органов ![]() , углерод
, углерод ![]() в слое почвы 0-30 см,
в слое почвы 0-30 см,
![]() и вынос углерода с сеном,
и вынос углерода с сеном, ![]() . Опыт продолжался
семь лет (табл. 3).
. Опыт продолжался
семь лет (табл. 3).
| г С/м |
г С/м |
|||||||
| Варианты опыта |
|
|||||||
| N-0 | 139 | 1232 | 12490 | 4731 | 923 | 3808 | 0 | 0 |
| N-1 | 200 | 1478 | 12400 | 5730 | 1048 | 3465 | 217 | 3.8 |
| N-2 | 247 | 1526 | 12915 | 8730 | 2710 | 5193 | 827 | 9.5 |
Внесение удобрений привело к повышению ![]() по сравнению с контролем в 1,2
раза на варианте N-1 (100 кг N/га) и в 1,8 раза на варианте N-2 (200 кг
N/га). Однако, наряду с
по сравнению с контролем в 1,2
раза на варианте N-1 (100 кг N/га) и в 1,8 раза на варианте N-2 (200 кг
N/га). Однако, наряду с ![]() увеличился и выходной поток
увеличился и выходной поток ![]() (вынос с сеном). В результате поступление растительных остатков
в почву несколько снизилось по сравнению с контролем на варианте N-1 и
значительно увеличилось на варианте N-2. Экосистема N-1 откликнулась на
изменение режима круговорота накоплением запасов растительного вещества
(вынос с сеном). В результате поступление растительных остатков
в почву несколько снизилось по сравнению с контролем на варианте N-1 и
значительно увеличилось на варианте N-2. Экосистема N-1 откликнулась на
изменение режима круговорота накоплением запасов растительного вещества
![]() и небольшим уменьшением запаса
и небольшим уменьшением запаса ![]() . В экосистеме N-2 произошло
увеличение как запасов растительного вещества, так и
. В экосистеме N-2 произошло
увеличение как запасов растительного вещества, так и ![]() . В обоих случаях
выполнялись условия переходного режима с накоплением:
. В обоих случаях
выполнялись условия переходного режима с накоплением:
![]() . Однако разница
. Однако разница
![]() составила всего лишь 3,8% от
составила всего лишь 3,8% от ![]() на варианте N-1 и значительно больше
(9,5% от
на варианте N-1 и значительно больше
(9,5% от ![]() ) на варианте N-2.
) на варианте N-2.
С течением времени переходной режим сменится периодическим и экосистема достигнет нового стационарного состояния с более высоким по сравнению с исходным общим запасом органического углерода. Прекращение удобрения почвы азотом переведет круговорот в переходной режим с потерями и экосистема вернется к исходному состоянию.
Дальнейшие наши работы выявили существование четвертого режима -- смешанного.
Смешанный режим характеризуется тем, что одни подсистемы биологического круговорота функционируют в периодическом режиме, а другие - в переходном. В таком случае в многолетнем разрезе для первых будут выполняться условия стационарности, для вторых - условия переходного режима с накоплением или потерями.
Исследование сукцессий и формирования БК позволили наметить четыре фазы его развития в травяных экосистемах.
Первая фаза начинается с момента включения фотосинтеза. Далее следует отмирание части
фитомассы автотрофов и минерализация растительных остатков. В определенное
время величина ![]() достигает терминального уровня и становится равной
разложению растительных остатков
достигает терминального уровня и становится равной
разложению растительных остатков ![]() , включающему минерализацию мертвой
фитомассы и образование почвенного органического вещества
, включающему минерализацию мертвой
фитомассы и образование почвенного органического вещества ![]() . Первая фаза
длится около 25 лет. За это время подсистема ``продуцирование
. Первая фаза
длится около 25 лет. За это время подсистема ``продуцирование ![]() фитомасса
фитомасса ![]() отмирание
отмирание ![]() растительные остатки
растительные остатки ![]() разложение
разложение
![]() '' выходит на стационарный уровень и начинает функционировать в
периодическом режиме.
'' выходит на стационарный уровень и начинает функционировать в
периодическом режиме.
Вторая фаза длится до выхода запасов биомассы почвенных микроорганизмов на стационарный уровень и захватывает период после 25 до 50 лет. После окончания второй фазы автотрофная и гетеротрофная подсистемы работают в периодическом режиме.
Третья фаза связана с накоплением почвенного органического вещества в аккумулятивном
горизонте почвы. Интенсивность образования ![]() растет по мере повышения
растет по мере повышения ![]() в
первой фазе и становится постоянной в течение второй фазы, а интенсивность
минерализации
в
первой фазе и становится постоянной в течение второй фазы, а интенсивность
минерализации ![]() увеличивается пропорционально растущему запасу
увеличивается пропорционально растущему запасу ![]() . Третья
фаза формирования БК, когда запасы всех компонентов, кроме
. Третья
фаза формирования БК, когда запасы всех компонентов, кроме ![]() , уже находятся
на стационарном уровне, а запас
, уже находятся
на стационарном уровне, а запас ![]() в верхнем слое (0-20, 0-30 см) почвы
продолжает накапливаться, длится около 100-150 лет.
в верхнем слое (0-20, 0-30 см) почвы
продолжает накапливаться, длится около 100-150 лет.
Четвертая фаза состоит в формировании полного профиля ![]() при стационарном состоянии
процессов продукции -- минерализации и структуры органического вещества в
верхних слоях почвы. Продолжительность фазы для разных почв -- от сотен до
тысячи лет.
при стационарном состоянии
процессов продукции -- минерализации и структуры органического вещества в
верхних слоях почвы. Продолжительность фазы для разных почв -- от сотен до
тысячи лет.
Каждая из этих четырех фаз является определенным этапом формирования БК. Вплоть до выхода всех подсистем БК на стационарный уровень, режим функционирования экосистемы -- смешанный.
 |
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Monday, 24-Sep-2001 22:28:39 NOVST